
Морфология и метаболизм дрожжей
Среди огромного разнообразия микроорганизмов, населяющих различные горизонты почв, обычно присутствуют и дрожжи. В то же время, численность дрожжей в почве невелика. Главным фактором, определяющим развитие дрожжей в почве, является концентрация легкодоступного органического вещества. Во всех типах почв численность дрожжей резко падает с глубиной, и наиболее заселенным является верхний 1-5см слой, непосредственно под подстилкой или слоем растительного опада.
Дрожжи играют большую роль в природных экосистемах. В трофических цепях эти дрожжи выступают как важное звено в питании беспозвоночных. Осуществляют процессы деструкции растительных остатков (Бабьева, Чернов, 2004).
В последние десятилетия разнообразие биотехнологических процессов, в которых используются дрожжи, резко увеличилось. Еще более разнообразны перспективы использования дрожжей: в различных разработках, патентах и т.п. упоминается более 200 видов. Сейчас дрожжи используются для получения различных ферментных препаратов, органических кислот, полисахаридов, многоатомных спиртов, витаминов и витаминных добавок, а также во множестве других мелкомасштабных процессах.
Целью исследовательской работы является выделение чистых культур почвенных дрожжей и создание питательной среды для культивирования базидиальных грибов на основе культуральной жидкости дрожжей.
Для достижения поставленной цели необходимо решить ряд задач:
- выделение чистых культур дрожжей и изучение морфологических и культуральных свойств;
- получение культуральной жидкости различных видов дрожжей и подбор питательной среды для культивирования высших грибов.
Глава 1. Морфология и метаболизм дрожжей1. Морфология
Для характеристик дрожжевых грибов обычно используют макро- и микроморфологические признаки, т.к. первые изучают визуально культуральные признаки, а вторые – с помощью микроскопа.
1.1 Макроморфология
Характеризует рост культуры в жидких или на плотных средах. Эти признаки очень изменчивы и сильно зависят от состава среды и условий культивирования, поэтому они имеют весьма ограниченное значение в систематике дрожжей. Но тем немее, многие виды дрожжей существенно отличаются по характеру роста на скошенном агаре или в виде гигантской колонии, что дает возможность их быстрой предварительной идентификации.
Дрожжевые культуры, растущие на плотных средах, по консистенции бывают чаще всего пастообразными, или стекающими полностью на дно пробирки, слизистыми, вязкими, клейкими, кожистыми или крошащимися. Слизистый рост характерен для многих анаморфных базидиомицетовых дрожжей родов Cryptococcus, Rhodotorula, Sporobolomyces, а также для аскомицетовых почвенных дрожжей рода Lipomyces. У большинства аскомицетовых дрожжей колонии пастообразные, сухие, культура при росте на скошенном агаре не стекает на дно пробирки. Для дрожжеподобных грибов, образующих как одиночные клетки, так мицелий, характеры колонии с ворсинчатым краем, который хорошо просматривается при просвечивании.
У большинства дрожжей колонии белые, часто приобретающие при старении кремовый или слегка коричневатый оттенок. У некоторых аскоспоровых дрожжей, например, из родов Nadsoniaили Lipomyces, старые колонии при обильном спорообразовании темнеют и становятся бурыми или шоколадными. Многие дрожжи образуют пигменты, окрашивающие колонии в разные цвета. Наличие каротиноидных пигментов, придающих колониям красную, розовую, оранжевую или желтую окраску, характерно для базидиомицетовых дрожжей родов Rhodotorula, Sporobolomyces, Cystofilobasidiumи др. Так же существуют дрожжи выделяющие в среду красно-вишневый пигмент пульхерримин. «Черные дрожжи» формируют темно бурые или черные колонии за счет накопления меланиноподобных пигментов.
При росте в жидких средах дрожжи вызывают помутнение, образуют осадок, кольцо на стенках пробирки, разного характера пленки. Как правило, рост в виде пленки характеризует способность дрожжей объединяться в мицелиальные структуры или свидетельствует об окислительном типе энергетического метаболизма (Бабьева, Чернов, 2004).
1.2 Микроморфология
Включает признаки, характеризующие отдельные клетки (форма, размеры), а также способы вегетативного и бесполого размножения и образуемые при этом структуры.
Для большинства аскомицетовых дрожжей характерно многостороннее почкование, почка возникает на любом месте клеточной поверхности последовательно одна за другой, но у некоторых видов может закладываться одновременно несколько почек. В этом случае речь уже идет о множественном почковании. При полярном почковании закладка почек происходит только строго по полюсам клетки и каждая последующая почка возникает на месте предыдущей. При этом шрамы почкования нарастают один на другой и придают клетке характерную форму: грушевидную (у рода Schizoblastosporion) или лимонную, апикулятную (у родов Saccharomycodes, Nadsonia и др.).
1.3 Псевдомицелий
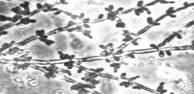
У многих видов дрожжей в определенных условиях роста материнские и дочерние клетки после почкования не разъединяются, а продолжают почковаться. В результате возникают структуры, имитирующие мицелий. Такой мицелий называют ложным, или псевдомицелием. В отличие от истинного(септированного) мицелия, в нитях псевдомицелия между клетками обычно хорошо заметны перетяжки, а апикальные (концевые) клетки всегда короче предшествующих. Псевдомицелий, стоящий только из клеток одного типа, сходных по форме и размерам, называют примитивным (рудиментарным). Сложный псевдомицелий состоит из клеток более чем одного типа, обычно в нем резко различают длинные клетки, составляющие псевдогифы, и расположенные на них одиночные или собранные гроздьями круглые, овальные или клиновидные почки, которые в этом случае называются бластоспорами (рисунок №1). Образование псевдомицелия характерно для многих аскомицетовых дрожжей, например, из родов Candida, Pichia (http://www.ss.msu.ru/soilyeasts/PicsList.htm)
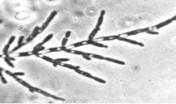
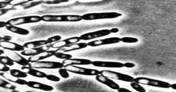
Рисунок 1. Примитивный - состоит из клеток одного типа

Сложный - состоит из клеток нескольких типов, обычно из удлиненных стволовых (псевдогифы) и более округлых мелких (бластоспоры).
1.4 Диморфизм и плеоморфизм
Для дрожжей, как и для других грибов, известны явления диморфизма и плеоморфизма. Мицелиально-дрожжевой диморфизм проявляется в том, что один вид может расти в двух формах – одноклеточной и мицелиальной. Это явление известно в микологии и связано с двумя типами роста – сферическим и апикальным. Есть дрожжи, которые образуют только одноклеточные популяции, хотя могут расти в виде конгломератов из отдельных клеток; они могут формировать структуры, внешне имитирующие мицелий. Типичный дрожжевой рост в процессе всего жизненного цикла характерен для многих видов аскоспоровых дрожжей. Другая группа дрожжеподобных грибов имеет либо мицелиальную, либо одноклеточную форму роста в зависимости от стадии жизненного цикла. Например, у базадомицетовых дрожжей обычно гаплоидная фаза одноклеточная, а диплоидная (дикариотическая) – мицелиальная.
Третья группа дрожжей проявляет дрожжевой или мицелиальный рост в зависимости от условий среды. Например, переход от одной клеточной фазе роста может быть связан с условиями аэрации.
О явлении плеоморфизма говорят, когда в жизненном цикле одного вида существуют два или несколько видов бесполого размножения. Плеоморфизм у дрожжеподобных грибов выражается в том, что наряду с основным типом вегетативного размножения почкованием или делением некоторые дрожжи образуют особые бесполые структуры, предназначенные специально для распространения или сохранения вида, например, баллитоспоры, эндоспоры, хламидоспоры.
Баллитоспоры – это экзогенные споры (конидии), которые формируются на заостренных кончиках особых выростов – стеригм, и при созревании с силой отстреливаются за счет капельно-экскреторного механизма. Способность к образованию баллитоспор, рассеиваемых через воздушную среду, свойственна дрожжам, обитающим на надземных частях растений. Примерами являются дрожжи родов Sporidiobolus, Sporobolomyces, Bullera и др.
Эндоспоры представляют собой бесполые эндогенные клетки, формирующиеся чаще всего в гифах мицелиальных дрожжей (например, Trichosporon) из участков цитоплазмы, которые отделяются мембраной и затем образую клеточную стенку. Количество эндоспор в одной клетке строго не фиксировано. После разрушения мицелия эндоспоры освобождаются и начинают почковаться. Эндоспоры ни по структуре, ни по устойчивости не отличаются от вегетативных клеток.
Хламидоспоры – крупные сферические или овальные клетки, которые образуются как из одиночных дрожжевых вегетативных клеток, так и на мицелии: интекалярно, латерально или терминально, по одной или цепочками. Хламидоспоры на мицелии образует, например, Candida albicans (рисунок №2). В хламидоспорах значительно снижена метаболическая активность, их клеточные стенки обладают устойчивостью к действию литических факторов. Биологическая функция таких структур заключается в длительном сохранении жизнеспособности в условиях голодания или низкой влажности. (http://www.ss.msu.ru/soilyeasts/PicsList.htm)
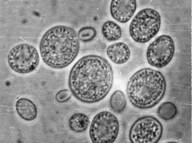
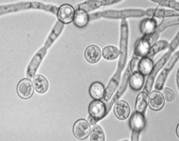
Рисунок 2. Phaffia rhorozyma Candida albicans
Истинный мицелий дрожжеподобных грибов имеет разное строение в зависимости от таксономического положения.
1.5 Особенности метаболизма
Хотя дрожжи и не так разнообразны по своему метаболизму, как бактерии, различные виды дрожжей могут катаболизировать разные соединения углерода и азота и образовывать разные конечные продукты
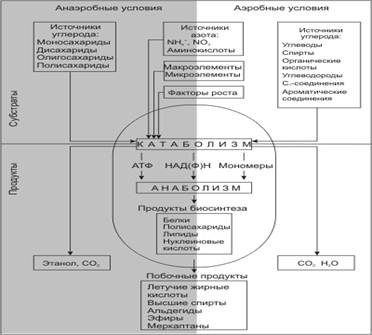
Рисунок 3. (http://www.ss.msu.ru/soilyeasts/PicsList.htm.)
1.5.1 Спиртовое брожение
Наиболее известное свойство многих дрожжей – способность к спиртовому брожению. Многие виды дрожжей могут переключаться с бродильного метаболизма на дыхательный и обратно в зависимости от условий: при наличии кислорода брожение ингибируется и дрожжи начинают дышать, в отсутствии кислорода включается механизм спиртового брожения. Так как кислородное дыхание – энергетически более выгодный процесс, чем брожение, то выход биомассы дрожжей в расчете на единицу используемого субстрата выше при выращивании их в аэробных условиях, чем в анаэробных. Это явление называется эффектом Пастера.
Спиртовое брожение может идти не только в анаэробных условиях. Если выращивать дрожжи в присутствии кислорода, но при высоком содержании глюкозы в среде, то в этом случае дрожжи также сбраживают глюкозу. Таким образом, глюкоза подавляет процессы анаэробного дыхания. Это явление получило название эффекта Кребтри, или катаболитной репрессии.
Многие дрожжи вообще не способны бродить. По соотношению между этими двумя процессами в метаболизме можно выделить следующие группы дрожжей.
1. Дрожжи, существующие только за счет брожения и не способные расти в аэробных условиях. К ним относятся, например, вид Arxiozyma telluris, обитающие в кишечном тракте грызунов.
2. Активные бродильщики интенсивно сбраживающие различные субстраты, но в анаэробных условиях переключаются на дыхательный обмен. Представители – Saccharomysec cerevisiae, Schizosaccharomyces pombe.
3. Слабые бродильщики в основном существующие за счет аэробного дыхания, но в аэробных условиях могут бродить, однако значительно менее интенсивно, чем виды из предыдущей группы. Это аскомицетовые дрожжи из родов Pichia, Dedaryomycas, а также все способные к брожению базидиомицетовые дрожжи.
4. Дрожжи существующие только а счет дыхания и не способные расти в анаэробных условиях. К этой группе относятся аскомицетовые дрожжи базидиомицетового аффинитета – Cryptococcus, Rhodotorula, Sporobolomyces.
Основными продуктами спиртового брожения являются этанол и углекислота, однако, в микроколичествах образуется также множество побочных соединений.
Субстраты брожения. Все бродящие дрожжи сбраживают глюкозу и фруктозу, поскольку именно с этих сахаров начинается гликолитическое расщепление. Кроме глюкозы и фруктозы могут сбраживать другие соединения, которые легко превращаются в интермедиаты гликолитического пути. В основном к ним относятся галактоза, из дисахаридов – сахароза, мальтоза, трегалоза. Значительно реже встречаются дрожжи, сбраживающие лактозу и мелибиозу.
Некоторые дрожжи способны сбраживать полисахариды. Например, Sacchoromycopsis fibuliger активно сбраживает крахмал.
Долгое время не были известны дрожжи, способные интенсивно сбраживать пентозы. Такие виды были описаны только к началу 80-х гг. XXв. К ним относятся, например, Pichia stipis (несовершенная стадия – Candida shehatae), Pachysolen tannophilus (Бабьева, Чернов, 2004).
1.5.2 Дыхание
При росте в аэробных условиях при низком содержании глюкозы в среде дрожжи получают АТФ за счет процессов дыхания, как это делает большинство аэробных организмов. Полное окисление субстрата до углекислого газа и воды может происходить у дрожжей с помощью трех различных механизмов: в цикле трикарбоновых кислот, в глиоксилатном цикле и в пентозафосфатном цикле.
Спектр углеродных соединений, усваиваемых дрожжами за счет аэробного дыхания, значительно шире, чем в случае брожения.
1.5.3 Вторичные продукты метаболизма
В процессе роста дрожжи на ряду с основными продуктами (этанол, СО2) образуют множество других метаболитов, которые при катаболизме углеводов, обмене аминокислот и пр. Хотя соединения обычно накапливаются в культуральной среде в очень незначительных количествах и трудно идентифицируются, они могут иметь большое практическое значение, например, для пищевой промышленности, так как от их состава зависит качество пищевого продукта, получаемого с помощью дрожжей. Такие соединения называют органолептическими.
Обязательные побочные продукты метаболизма дрожжей – высшие спирты (сивушные масла). Механизм синтеза высших спиртов связан с образованием алифатических аминокислот.
Чаще всего встречаются спирты: пропиловый, изоамиловый, бутиловый, изобутиловый. Летучие жирные кислоты, такие как уксуная, пропионовая, масляная, изомасляная, изовалериановая – также обычно минорные продукты метаболизма дрожжей. Большое практическое значение имеют выделяемые дрожжами альдегиды и кетоны (ацетоин и диацетил) (Бабьева, Чернов, 2004).
1.6 Источники азота
Универсальным источником азота для дрожжей являются соли аммония.
Многие дрожжи способны усваивать азот в окисленной форме – в виде солей нитратов и нитритов. Дрожжи, использующие нитраты, имеют две ферментные системы: первая восстанавливает нитрат до нитрита, вторая – нитрат до аммония. У некоторых видов присутствует только вторая ферментная система – они способны усваивать нитриты, но не нитраты. Способность к ассимиляции нитратов считается ценным таксономическим признаком, поэтому определение способности к росту на среде с KNO3 в качестве единственного источника азота – рутинный тест при идентификации дрожжей.
Практически все дрожжи используют в качестве источника азота мочевину, расщепляя ее до СО2 и NH3.
1.7 Лимитирующие факторы
За исключением нескольких исключительно холоднолюбивых видов, среди дрожжей нет ярко выраженных экстремофилов, то есть видов, предпочитающих крайне высокие или низкие значения температуры, рН, осмотического давления, влажности, среды, и т.п. В тоже время существуют дрожжи, которые сильно выделяются среди большинства других видов, которые сильно выделяются среди большинства других видов по своей способности переносить неблагоприятные факторы среды (http://www.ss.msu.ru/soilyeasts/PicsList.htm.).
2.1 Объекты исследований
Объектом исследований является почва, отобранная из различных экониш. Проба №1 – почва поля не посевного (Ахтубинский р-он), проба №2 – почва лесная (Ахтубинский р-он), проба №3 – почва садовая (парк АГТУ), проба №4 – почва луговая (луг заливной, Ахтубинский р-он), проба №5 – почва из грибницы шампиньона (Икряненский р-он).
2.2 Отбор проб
Среднюю почвенную пробу получают смешиванием отдельных образцов. Пробы №1, №2, №3, и №4 были отобраны по одном и тому же методу, с площади 100 м2 взяли пробу из трех точек. Верхний слой почвы толщиной 2 см снимается и только после этого отбирается проба почвы. Так проделываем три раза. Проба №5 отбирается из одной точки, из грибницы шампиньона.
2.3 Приборы для взятия проб
Почвенный образец берут буром, лопатой и ножом. В поле перед взятием образца их тщательно очищают, затем обрабатывают спиртом и обжигают. Можно ограничиться очисткой этих предметов, если затем несколько раз воткнуть их в почву изучаемого горизонта. Укладывают образец в заранее приготовленную стеклянную широкогорлую стерильную банку, закрывающуюся корковой пробкой, обернутой стерильной бумагой, либо в стерильные пергаментные пакеты или пакеты из плотной бумаги, взятой двойным слоем. На пакеты, банки наклеивают этикетки с указанием места взятия пробы, горизонта и других сведений.
Почвенные образцы анализируют в первые сутки. При необходимости допускается хранение их в холодном помещении (в холодильнике) в течение двух дней. Для придания среднему образцу большей однородности, соблюдая все условия асептики, тщательно перемешивают почву, вынимают корни растений, различные включения (Теппер, 1994).
2.4 Методы стерилизации
Стерилизация, или обеспложивание (от лат. sterilis – бесплодный), – это полное уничтожение клеток микроорганизмов в питательных средах, посуде и пр.
Известно несколько методов стерилизации. Чаще всего применяют стерилизацию нагреванием.
Фламбирование, или прокаливание
Прокаливать можно непосредственно перед употреблением платиновые петли, иглы, шпатели, мелкие металлические предметы (ножницы, ланцеты, пинцеты), а также стеклянные палочки, предметные, покровные стекла и т. д.
Стерилизация сухим жаром
Ее применяют для обработки посуды и сухих материалов, например крахмала, мела. При этом стерилизуемый объект выдерживают при 170ºС в течение 2 ч (считая с того момента, как установлена необходимая температура) в печи Пастера или в электросушильных шкафах. Поднимать температуру выше 170ºС не рекомендуется: ватные пробки и бумага начинают разрушаться (буреют, становятся ломкими).
Перед стерилизацией стеклянную посуду закрывают ватными пробками и обертывают бумагой. Чашки, пробирки, пипетки, вату, марлю заворачивают в бумагу или помещают в особые футляры и пеналы, в которых стерильная посуда может храниться после стерилизации.
По окончании стерилизации шкаф открывают только после того, как температура снизится до комнатной, иначе стекло может лопнуть.
Стерилизация текучим паром
Текучим паром (100 °С) обрабатывают предметы, портящиеся от сухого жара, и некоторые питательные среды, не выдерживающие более высокой температуры (среды с углеводами, МПЖ, молоко). Проводят стерилизацию в кипятильнике Коха по 30 мин в течение 3 суток ежедневно. Такая стерилизация называется дробной.
Стерилизация насыщенным паром под давлением
Это наиболее быстрый и надежный способ стерилизации, при котором гибнут самые устойчивые споры. С его помощью стерилизуют большинство питательных сред, посуду.
Обработку насыщенным паром проводят в герметически закрывающемся толстостенном котле – автоклаве. На массивной крышке или сбоку котла находятся кран для выхода пара, манометр и предохранительный клапан. Манометр показывает, на сколько давление пара внутри котла выше нормального. Для предотвращения взрыва при превышении предельного давления срабатывает предохранительный клапан, давая выход пару.
Автоклав используют и для дробной стерилизации текучим паром. В этом случае крышку не завинчивают, чтобы обеспечить свободный выход пару (Теппер, 1994).
2.5 Материалы и оборудование
Культуры микроорганизмов, пробирки, колбы, чашки Петри, бактериологические иглы, шпатели, штативы, предметные и покровные стекла, красители, спиртовая горелка.
При любой микробиологической работе: при посевах, при посевах, выделении, пересевах, сохранении чистых культур используются стерильные среды, стерильная посуда, стерильные инструменты, чтобы предотвратить возможность попадания посторонней микрофлоры.
Для стерильной посуды используется стерилизация сухим жаром. Она производится нагреванием в течение 2 часов, при температуре 1700°С (считая с того момента, как температура установилась) в печи Пастера или в электросушильных шкафах (с нагревом до 2000).
Вся посуда перед стерилизацией должна быть вымыта, высушена и завернута в бумагу.
Для заворачивания посуды при стерилизации в ав