Структура биологической мембраны
(современная концептуальная модель биомембраны Сингера-Николсона[НД23] )[Мф24]
Клеточные мембраны построены по общей схеме. Основой всех биомембран является двойной слой липидов (фосфолипидов и гликолипидов)[Мф25] .
Почему эти липиды стали основой всех клеточных мембран? Находясь в водной среде, они обладают свойством самоорганизовываться. Как? Эти липиды амфифильны, т.е. имеют гидрофильную (полярную) и гидрофобную (неполярную) части [5] (рис. 409122014).

Рис. 409122014. Гидрофильная [V.G.26] часть (головка) (5) фосфолипида представлена фосфатным остатком (2) с присоединённым к нему холином (1), этаноламином или серином. Гидрофобную часть (хвосты) (4) составляют жирные кислоты.[6]
| Гидрофобные хвосты поворачиваются друг к другу, а гидрофильные головки соприкасаются с водными фазами. | 
|
| Причём образуются замкнутые структуры клетки, другими словами они ограничивают некоторый объём (полость мембраны) от внешней среды или других частей клетки.[7] | 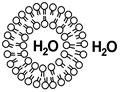 [8] [8]
|
| Ядро и митохондрии имеют двойную мембрану, т.е. двойной бислой фосфолипидов [9] → | 
|
Включённые в мембрану белки, как правило, делят на интегральные (синоним – внутренние [10]) и периферические (синонимы: поверхностные, наружные [11]), часто выделяют полуинтегральные белки (рис. 409111002).

| Рис. 409111002. Белки биомембраны: 1 – интегральный, 2 – периферические, 3 – полуинтегральный. |
Итак, в общем плане, биомембраны состоят из бислоя фосфолипидов и включённых в них белков. А теперь несколько подробнее о строении биомембран.
Прежде всего, следует отметить, что внешняя и внутренняя поверхность мембран различаются по составу липидов, белков и наличию углеводов — поперечная асимметрия. [12]
Мембраны включают в себя много холестерина (рис. 409122241):[13]

| Рис. 409122241. Включение молекул холестерина в биомембрану (обозначены Х): [14] |
Различия периферических и интегральных белков определяют степень связывания их с мембраной, но не способ их прикрепления к бислою.[15]
Способы прикрепления белков к мембране разнообразны:

| Связывание с "якорными" белками, погруженными в бислой. Примеры: F1‑часть Н+‑АТФазы связана с F0‑частью, погруженной в мембрану; сукцинатдегидрогеназа, некоторые белки цитоскелета. |
| Связывание с поверхностью бислоя | |
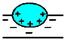
| электростатической природы. Пример: миелиновый основный белок. |
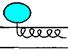
| гидрофобной природы, но практически без погружения в бислой. Пример: пируватоксидаза, фосфолипазы. |
| Связывание с помощью гидрофобного "якоря" | |
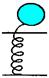
| Пример: Цитохром b5 имеет короткий концевой сегмент из неполярных аминокислотных остатков. |
| Некоторые белки используют в качестве "якоря" ковалентно связанные с ними жирные кислоты или фосфолипиды, например, щелочная фосфатаза эукариот |
| Пересечение мембраны трансмембранными белками | |
| Гликофорин имеет одиночный трансмембранный сегмент |
| Лактопермеаза и бактериородопсин имеют несколько трансмембранных сегментов. |
Ещё раз рассмотрим схему локализации белков в мембране из учебника [b] с которой Вы подробно будете знакомиться на последующих занятиях и лекциях (рис. 40912213).
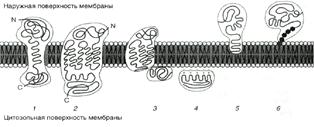
Рис. 40912213. Локализация белков в мембранах. 1,2— трансмембранные белки, пример: гликофорин, рецептор адреналина; 3— связывание с белками, погруженными в бислой, пример: фермент митохондрий — сукцинатдегидрогеназа; 4— связывание с поверхностью бислоя, пример: миелиновый основной белок; 5— «заякоривание» с помощью короткого концевого домена, пример: цитохром b5 ; 6— «заякоривание» с помощью ковалентно-связанного липида, пример: фермент щелочная фосфатаза.
| Интегральные белки образуют гидрофильные каналы → Подробнее смотри далее или [--144-С.98] | 
|
Приведём некоторые схемы, показывающие более подробно строение биомембраны (рис. 409122245, 409122246, 409122247).
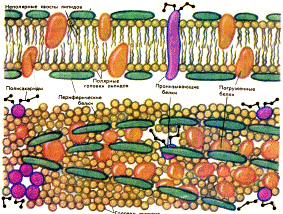
Рис. 409122245. Схема строения цитоплазматической мембраны: внизу — вид сверху, .вверху — поперечный разрез [16]

Рис. 409122246. Трехмерная схема жидкостно-мозаичной модели мембраны[17]
1 — гликолипид, 2 — ионный канал, 3 — фосфолипид, 4 — интегральный белок, 5 — олигосахаридная боковая цепь, 6 — гидрофобный участок а-спирали, 7 — а-спиральная белковая молекула, 8 — холестерин, 9 — наружная поверхность, 10 — липидная сердцевина, 11 — внутренняя поверхность.
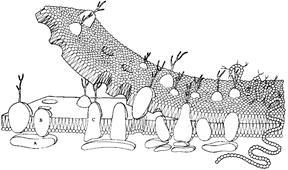
Рис. 409122247. Трехмерная модель биомембраны с с частичным разделением бислоя.[18]
Биологическая мембрана часто является частью оболочки (рис. 409122254).
Клеточная оболочка (поверхностный аппарат) включает три компонента: плазматическую мембрану (плазмолемму), надмембранный и подмембранный слои[Б27] . Коротко остановимся на околомембранных слоях, подробно см. [++598+, С.6-9].
Надмембранный слой (гликокаликс) имеет нежную фибриллярную структуру [Б28] . В его состав входят периферические белки и углеводные компоненты гликолипидов и гликопротеинов плазмолеммы, частично интегральных белков (см. рис[Б29] . ), а также различные соединения, адсорбированные клеточной поверхностью[Б30] . Существенно, что в разных клетках организма интенсивность развития надмембранного слоя, его химический состав, а также функции неодинаковы[Б31] [Б32] . Толщина гликокаликса – порядка 50 нм [Б33] , т.е. он в 5 раз толще мембраны.
Подмембранный слой, представленный узким участком цитоплазмы, примыкает к плазмолемме с внутренней стороны[Б34] . Гиалоплазма в этой области более вязкая и практически не содержит органелл[Б35] . Здесь сосредоточены структурные элементы опорно-сократительного аппарата клетки — цитоскелета[Б36] .

Рис. 409122254.[19] Схема оболочки клетки: A – надмембранный слой (гликокаликс); B – плазматическая мембрана; C – подмембранный слой.
1 – гликопротеид, 2 - гликолипид
На рисунках 409122308, 409122309, 409122310 разными авторами по-разному, но достаточно хорошо показана организация подмембранного слоя.

Рис. 409122308.[20] Схематическое изображение клеточной мембраны.

Рис. 409122309.[21] Схематическое изображение клеточной мембраны с элементами цитоскелета.
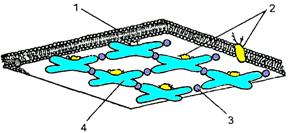
Рис. 409122310.[22] Схема организации спектрин-актиновой сети: 1 — липидныи бислой; 2 — интегральные белки; 3 — актин; 4 — тетрамер спектрина