Механизмы созревания и дифференцировки В- и Т-лимфоцитов
Лимфоидная система представлена первичными лимфоидными органами – костным мозгом и тимусом у млекопитающих – и множеством вторичных лимфоидных органов, в которых происходит антигеннезависимое и антигензависимое взаимодействиезрелых циркулирующих лимфоцитов. В большинстве случаев зрелые лимфоциты впервые контактируют с антигеномв лимфоидных органах; сенсибилизациялимфоидных клеток вне лимфоидных органов происходит весьма редко.
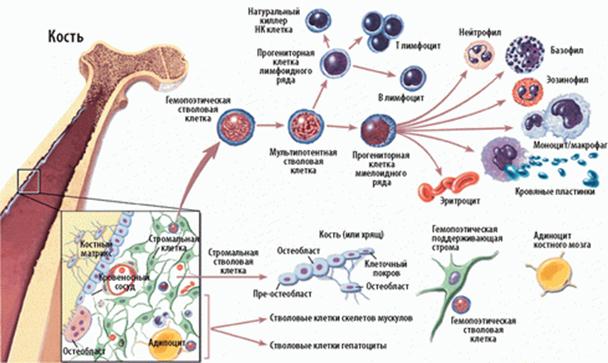
Считается, что лимфоциты, как и все остальные элементы крови, образуются из популяции изначальных гематолимфоидных предшественников, которые впервые обнаруживаются на переднем конце эмбрионов позвоночных. Затем гемопоэтические стволовые клетки появляются последовательно в кровяных островках желточного мешка, в эмбриональной печени, в эмбриональной селезенке (очень недолго) и, наконец, в костном мозге.
Частично дифференцированные стволовые клетки, запрогаммированные к лимфоидной дифференцировке, сначала обнаруживаются в желточном мешке, но со временем «заселяют» костный мозг.
Примечание: Клетку называют стволовой,если одна из возникающих в результате деления дочерних клеток дифференцируется, а другая остается недифференцированной и служит источником последующих поколений дифференцировонных клеток. Такое ассиметричное деление характерно не только для стволовых клеток. Клетки , сохранившие способность лишь к одному пути дифференцировки, также могут оставаться способными к самообновлению. Прямых экспериментальных доказательств существования общей лимфоидной стволовой клетки пока ещё нет.
|
У млекопитающих костный мозг представляет собой место созревания В-линии, превращающихся из стволовой клетки последовательно в большие пре-В-, малые пре-В- и малые зрелые В-лимфоциты. Тонкая структура той среды в костном мозге млекопитающих, в которой происходит созревание В-лимфоцитов, изучена ещё недостаточно. Костный мозг содержит гетерогенную популяцию нелимфоидных прикрепленных клеток, необходимых для созревания всех основных нелимфоидных родоначальных клеток. Пре-В-клетки костного мозга экспессируют поверхностные маркеры, присущие развивающимся В-клеткам. У мышей на них приходится не менее 30% костномозговых клеток, имеющих ядро, и большинство костномозговых лимфоцитов.
В больших пре-В-клетках впервые происходит перестройка генов, кодирующих тяжёлые (Н) цепи иммуноглобулинов ( в результате образуются функциональные транскрипты для m-цепей). Затем эти клетки делятся, давая начало малым пре-В-клеткам. В малых пре-В-клетках перестраиваются гены легких капа(æ)-цепей , что сопровождается появлением этих цепей в цитоплазме и экспрессией поверхностных IgMæ. У мышей очень небольшая часть В-клеток претерпевает дальнейшую перестройку генов легких l-цепей (95æ:5l) с последующей экспрессией этих цепей. У людей это происходит также, как правило, и приводит к частичной или полной делеции ранее сформированных æ-генов (70æ:30l).
Созревание В-клеток считается завершенным, когда на их поверхности экспрессируются иммуноглобулиновые рецепторы для антигена, структуры, кодируемые генами главного комплекса гистосовместимости (major hystocompatibility complex –MHC), маркеры иммунологической индивидуальности, необходимые для осуществления межклеточных взаимодействий при развитии иммунного ответа, и рецепторы, наличие которых позволяет В-клеткам поступать во вторичные лимфоидные органы.
Итак, В-лимфоциты берут начало из стволовых клеток костного мозга. Лимфоциты этого класса завершают своё развитие в костном мозге. Они сходны по размеру и внешнему виду с Т-клетками.
В-лимфоциты можно выявить с помощью специфических антисывороток против иммуноглобулинов (Ig). Оказалось, что
в тимусе – 0,2% этих клеток,
в грудном лимфатическом протоке – 15%,
в лимфатических узлах – 25%,
в периферической крови – 25-30%,
в селезенке – 40%,
в костном мозге – 40%,
в пейеровых бляшках –55%, почему их и относят к центральному органу иммунной системы, В-системы иммунитета у млекопитающих, аналогу сумки Фабрициуса у птиц.
В периферической крови 55-60% Т-лимфоциты, 25-30% - В-лимфоциты и 10-20% не обладают признаками ни Т-, ни В-лимфоцитов (нулевые).
В-лимфоциты – продуценты антител или иммуноглобулинов. Согласно клонально-селекционной теории, предложенной в 1959 году Ф.Бернетом, каждая В-клетка, завершив свое развитие в костном мозге становится продуцентом антител, узнающих один специфический антиген, то есть одну молекулярную структуру.
Антитела, синтезированные в В-клетке, остаются связанными с её клеточной мембраной, где они располагаются на поверхности в качестве молекул-рецепторов. Связывание антигена с антителом на мембране является для клетки стимулом к пролиферации – в этом состоит отбор (селекция) клонов.
Некоторые из клеток-потомков отобранных таким образом клонов остаются циркулировать в организме. Эти В-лимфоциты служат памятью иммунной системы: когда в организм попадает тот же антиген, они обеспечивают быстрый вторичный ответ. Такие клетки циркулируют в крови в течение многих месяцев и лет. Клетки памяти ответственны за иммунный ответ, который развивается после перенесенной инфекции или вакцинации.
Другие члены отобранных клонов претерпевают окончательную дифференцировку: они вырастают до больших размеров, прекращают размножение и интесивно продуцируют антитела. Такие В-клетки ( в этом состоянии их называют плазматическими клетками) живут только несколько дней, но успевают произвести огромное количество иммуноглобулинов.
Молекулы антител сами не могут разрушить чужеродный организм, они лишь отмечают его, чтобы его смогли узнать другие защитные системы, такие как система комлемента, макрофаги и другие клетки. Три молекулы антитела, связавшиеся с поверхностью бактерии и расположенные на ней близко друг к другу, притягиваются к макрофагу в тысячу раз сильнее, чем одиночная молекула антигена.

|
Две “триады” приобретенного иммунитета составляют едва ли не всю суть иммунологии: