ИММУНОГЕНЕТИКА
Наличие лимфоидных клеток, способных синтезировать антитела отдельных классов, подклассов, аллотипов и идиотипов, говорит о том, что механизмы создания разнообразия антител (специфичности) контролируются генетически.
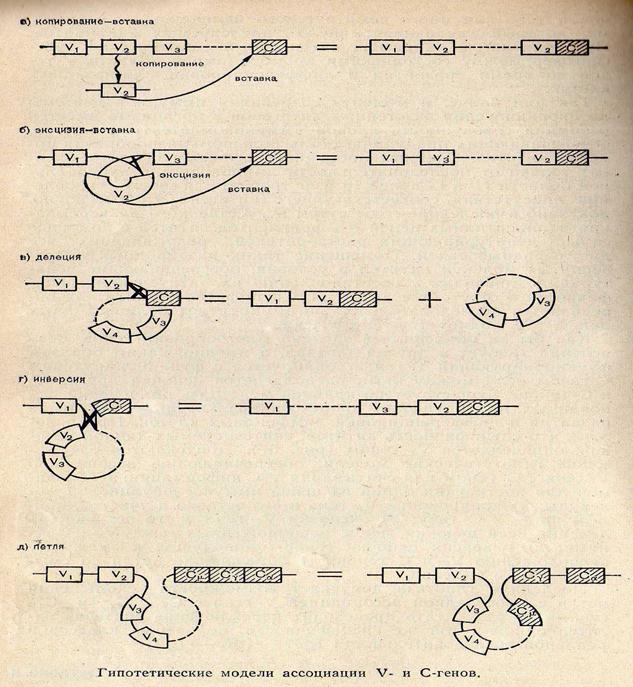
Применение методов молекулярной гибридизации позволило установить, что гаметные С-гены уникальны. Существует только один С-ген цепей æ, четыре гена цепей lи по одному гену для каждого класса и подкласса тяжелых цепей. Væ- и Vl-гены могут соединяться только со своими Сl-генами, образуя VæCæ- и VlCl -цистроны, в то время как Vн-гены могут соединяться с любым Сн-геном. Объединение Vн и Сн-генов приводит к образованию цистронов тяжелых цепей для каждого из классов: VmCm , VgCg, VaCa, VdCd, VeCe. C-гены и V-гены цепей æ и l у мышей располагаются на 6-й и 16-й хромосомах соответственно, гены же тяжелых цепей расположены на 12-й хромосоме, то есть оба гена легких цепей и гены тяжелых цепей располагаются на разных хромосомах, поэтому ни рекомбинации, ни кроссинговер между ними не происходит.
Специфика организации тяжелых генов состоит в том, что в процессе дифференциации антителообразующей клетки возможно последовательное переключение с Сm-гена на Сg- и Сa-гены, а также переключение мембранных форм иммуноглобулинов на секреторные, отличающиеся длиной хвостовых участков.
Как æ- и l-гены, гены тяжелых цепей построены из экзонов и интронов. Количество экзонов соответствует числу Сн-доменов, константный ген тяжелых цепей содержит 3-4 экзона и 2-3 интрона. Все Сн-гены в эмбриональных клетках не сцеплены, а располагаются отдельной группой на расстоянии нескольких тысяч пар оснований от Vн-генов. Сегменты ДНК между отдельными Сн-генами имеют длину несколько десятков тысяч пар оснований.
Таким образом, в настоящее время установлено, что каждая тяжелая и легкая полипептидная цепь иммуноглобулинов кодируется несколлькими генами, которые в ДНК первичных половых клеток физически разобщены, но объединяются в В-лимфоцитах и антителообразующих клетках, образуя единый активный ген ( то есть догма: один ген – один фермент, одна полипептидная цепь – не состоятельна для иммунной системы).
Вариабельные домены легких цепей æ-типа контролируются двумя разными генами Igæ-V(Væ) и Igæ-J(Jæ). У мыши Væ-ген кодирует первые 95 аминокислотных остатков, а Jæ- остальные 12 аминокислот.
.

| Впервичных половых клетках эти Væ- и Jæ-гены находятся на большом расстоянии друг от друга, но в ходе развития лимфоцита они объединяются. При ассоциации с геном константной части æ-цепи, расположенным сравнительно близко к Jæ-гену, получается единый активный ген. Количество Væ-генов, способных участвовать в формировании æ-цепей примерно 300, а Jæ-цепей – 5, хотя функционируют, вероятно, только 4. Соответственно комбинаторное объединение Væ- и Jæ-генов теоретически может обеспечить примерно 1200-1500 вариантов легких цепей. Ещё большее разнообразие может возникнуть в том случае, если транслокация, приводящая к объединению Væ- и Jæ-генов, происходит не совсем точно и в результате на стыке этих двух генов кодируются разные аминокислоты (ошибки сплайсинга) Количество легких цепей æ-типа, которые могут быть построены в результате комбинаторного объединения, может увеличиваться ещё больше в результате соматических мутаций (3 мутации на 30 делений (!), вместо одной мутации на миллион). Таким образом, при довольно ограниченном количестве генетической информации, имеющейся в первичных половых клетках (гаметах) число различных Læ-цепей может быть весьма большим. | |
| Гены тяжелых цепей организованы в принципе также, но обладают еще более сложной организацией. Вариабельный домен тяжелой цепи образуется благодаря комбинаторному объединению трех типов геметных генов Igh-V(Vн) - 350, Igh-D(Dн) - 20 и Igh-J(Jн) – 4. Это обеспечивает ещё большее разнообразие тяжелых цепей по сравнению с легкими цепями. Кроме того, один и тот же генный комплекс может быть экспрессирован вместе с любым из генов, кодирующим константные области тяжелых цепей (Igh-C-гены). | 
| |
| ||
Jн-гены расположены недалеко от Igh-m-гена, поэтому сборка генного комплекса VDJ происходит рядом с m-геном константной области. Этим объясняется тот факт, что в ходе иммунного ответа первым синтезируется именно IgM-антитела, и они же содержатся в мембране большинства В-лимфоцитов в качестве рецепторов.
У мыши константные участки тяжелых цепей расположены в следующем порядке, начиная с 5’-конца: m, d, g, e, a. Вероятно, в процессе транскрипции образуется единая первичная РНК, которая соответствует V,D,J-, m- и d-гену, а также большим отрезкам ДНК, разделяющим эти гены. Последующее формирование зрелой РНК для синтеза m- или d-цепей осуществляется путем сплайсинга. Такая схема хорошо объясняет совместную экспрессию IgM и IgD в качестве мембранных иммуноглобулинов на поверхности большинства клеток. Для возникновения генов, обеспечивающих синтез IgG, IgA, IgE требуется, по-видимому, какой-то другой генетический механизм.
Очевидно при этом происходит транслокация (перемещение) гена VDJ из положения 5’ по отношению к гену m в аналогичное положение 5’ по отношению к генам g, e и a. Действительно, на 5’ концах всех генов константной области (за исключением d) были обнаружены характерные последовательности, обеспечивающие транслокацию. Их обозначили как участки переключения (S-районы).
Таким образом, биологический феномен переключения обусловлен рекомбинациями, касающимися константных (С) областей иммуноглобулинов (Ig). Способ рекомбинации, его регуляция изучаются.
Терминологический словарь:
рекомбинация – изменение генетической карты;
транспозиция – изменение места расположения гена или его части;
транслокация – перемещение;
делеция – выпадение;
сплайсинг – сочленение, склеивние;
процессинг – созревание;
репликация – удвоение ДНК;
транскрипция – образование копии одной из цепей ДНК в виде РНК;
трансляция – биосинтез полипептидной цепи;
экзон – структурный участок гена;
интрон – разделительный, вставочный участок гена.