Метод локальной фиксации напряжения (пэтч-кламп).
Ионные каналы биологических мембран исследуют главным образом с помощью метода локальной фиксации напряжения, называемого также методом пэтч-кламп. В этом методе напряжение фиксируют на микроучастке клеточной поверхности, что позволяет регистрировать токи одиночных ионных каналов. Первые такие измерения, были осуществлены Э. Неером и Б. Сакманом в 1976 г. Этот метод позволяет регистрацию токов в микроучастках мембраны диаметром примерно 1 мкм и позволяет идентифицировать молекулярные реакции одиночных каналов на основе зависимостей ионных токов от потенциала и времени (слайд 4). Стеклянная микропипетка, диаметр кончика которой меньше 1 мкм, подводится к клетке вплоть до контакта с мембраной, и когда через пипетку подается отрицательное давление, пипетка обычно закупоривается участком мембраны; электрическое сопротивление между пипеткой и внеклеточным раствором возрастает скачком более чем до 5 ГОм (109 Ом).
В результате микроучасток мембраны электрически изолируется от остальной мембраны. Канал пипетки соединен с усилителем обратной связи, который обеспечивает регулирующую цепь для поддержания потенциала пипетки на заданном уровне. Ток, необходимый для стабилизации потенциала-«ток фиксации» - точно соответствует току, протекающему в каждый момент через микроучасток мембраны.
Благодаря механической прочности контакта между стеклом в торце микропипетки и клеточной мембраной мембранный фрагмент можно отделить от клетки, получив доступ к его внутренний поверхности, либо разрушить мембранную перегородку в кончике пипетки и получить сравнительно низкоомный контакт с внутриклеточной средой. В зависимости от условий различают несколько вариантов измерений: 1) на микроучастке мембраны с прикрепленной клеткой, 2) на изолированном участке мембраны с внешней поверхностью, обращенной внутрь пипетки, 3) на изолированном фрагменте, внешняя поверхность которого обращена наружу, 4) измерение интегральных токов на целой клетке (метод внутреннего диализа).
2.3. Ионные токи при возбуждении Временной ход Na+ и К+ - токов и изменений проводимости.
Деполяризация, возникающая во время возбуждения, изменяет проводимость мембраны для различных ионов, а сдвиг проводимости в свою очередь вызывает изменение потенциала.
В ответ на ступенчатую деполяризацию аксона кальмара от потенциала покоя до 0 мВ регистрируется мембранный ток, состоящий их 4х компонентов (слайд 2). Емкостной ток Jс представляет собой кратковременный импульс, который в основном заканчивается к моменту выхода напряжения на плато. Некоторые дипольные молекулы в мембране медленно меняют свою ориентацию после наложения ступенчатого напряжения, что приводит к появлению небольшого по амплитуде "хвоста" емкостного тока, который относительно медленно спадает. После спада Jс мембранный ток полностью определяется движением ионов через каналы в мембране.
В ионном токе можно выделить два основных компонента (Na+ и К+ токи) и один второстепенный – ток утечки Je.
Рассматривая основные компоненты: калиевый и натриевый токи, необходимо отметить, что натриевый ток обусловлен преимущественно кратковременным потоком Na+, проходящим через Na+-каналы. Этот ток быстро уменьшается и спустя примерно 1мс сменяется входящим К+- током.
Временной ход Na+ и К+ - токов и изменений
проводимости.
Ток фиксации J, возникающий в ответ на скачок деполяризации состоит таким образом из Na+ и К+ токов (JNa и JK) и зависит от мембранного потенциала. Эквивалентная схема, используемая при интерпретации данных в опытах по фиксации потенциалов предполагает, что токи, текущие по Na+ и К+ каналам подчиняются уравнениям:
 ,
, 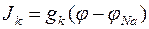 , (1).
, (1).
Если во время развития ионного тока скачком сместить мембранный потенциал на новый уровень, то ток тоже изменится скачком. Изменение тока Na+ и К+ во времени в режиме фиксации напряжения обусловлено изменением проводимостей gNa и gk поскольку величина 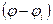 остается постоянной в течение всего импульса. При ступенчатой деполяризации мембраны Na+-проводимость начинает изменяться от чрезвычайно малой величины, быстро нарастает до 25 мОм/см2, а затем экспоненциально уменьшается (слайд 3).
остается постоянной в течение всего импульса. При ступенчатой деполяризации мембраны Na+-проводимость начинает изменяться от чрезвычайно малой величины, быстро нарастает до 25 мОм/см2, а затем экспоненциально уменьшается (слайд 3).
Проводимость для К+ нарастает по S-образной кривой и выходит на постоянный уровень через 5-6 мин. Изменения проницаемости для Na+ и К+ градуальны и обратимы. Натриевая проводимость восстанавливается примерно в 10 раз быстрее, чем калиевая.
Na-проводимость может быть уменьшена двумя разными способами. Если мембранный потенциал возвратить на исходный уровень через короткий промежуток времени, то система, регулирующая Na+-проницаемость, быстро возвращается к состоянию покоя. Второй импульс, нанесенный сразу же после первого, вызывает в этом случае повторное увеличение Na+-проводимости. Если же деполяризация продолжительна, Na+-проводимость уменьшается более медленно вследствие процесса, называемого инактивацией. Для того что бы второй импульс после инактивации Na+-каналов мог снова вызвать изменение проницаемости для Na+ , мембрана в течение нескольких миллисекунд до начала второго импульса должна быть реполяризована.