Figure 1 - Functional chart (diagram) of DTS TS with direct coding
Водоросли – это прокариотические и эукариотические талломные споровые организмы, содержащие, как правило, в своих клетках хлорофилл, обладающие оксигенным фотосинтезом, являющиеся аэробами и не достигшие уровня дифференциации, характерной для архегониальных растений.
| Водоросли | Высшие растения |
| Возникли раньше (архейская эра, 4 млрд лет). | Возникли позже (силур палеозойской эры). |
| Первичноводные. | Наземные и вторичноводные. |
| Произошли в разное время от разных предков. Распределены среди 4 царств (Сyanobiontes, Euglenobiontes, Chromista, Plantae). | Произошли от водорослевого предка и относятся к 1 царству Plantae. |
| Различные типы питания: автотрофное, гетеротрофное, миксотрофное (включая голозойное). | Преимущественно автотрофное питание, реже вторично гетеротрофное, либо авто-гетеротрофное. |
| Различный уровень организации (одноклеточный, колониальный, многоклеточный). | Только многоклеточный уровень организации. |
| Талломные (слоевищные) организмы. | Листостебельные, реже талломные, организмы. |
| Разнообразие морфоструктуры таллома (монадный, амебоидный, пальмеллоидный, коккоидный, нитчатый, разнонитчатый, паренхиматозный, псевдопаренхиматозный, харофитный, сифональный, сифонокладальный) (см. ниже). | У талломных только паренхиматозный (тканевой) таллом. |
| Ткани только у водорослей с паренхиматозным талломом (часть зеленых, бурых, красных, желтозеленых, золотистых). Из тканей только паренхима, реже (бурые) покровная, образовательная, проводящая (“ситовидные трубки”), механическая. | Ткани у всех. Различные ткани. |
| У большинства отсутствуют механические структуры. Есть только у бурых в сердцевине). Вода – плотнее воздуха. | Имеются механические ткани. |
| У подавляющего большинства нет проводящих тканей. | Есть проводящие ткани. |
| Нет устьиц, газообмен всей поверхностью тела. | Есть устьица. |
| У большинства нет покровных тканей. | Есть покровные ткани. |
| Всасывание минеральных веществ всей поверхностью тела. Ризоиды только для прикрепления. | Всасывание воды и минеральных веществ с помощью корневых волосков у большинства. |
| Покровы клеток разнообразны (цитоплазматическая мембрана, пелликула, перипласт, тека, панцирь, домик, клеточная оболочка). | Клетки имеют клеточную оболочку. |
| Клеточная стенка имеет разнообразный состав: целлюлоза, каллоза, муреин, пектин, углекислая известь, альгиновая кислота, фикоколлоиды. Лигнина нет. | Клеточная оболочка из целлюлозы, гемицеллюлозы, пектина. Могут откладываться лигнин, суберин. |
| Прокариоты и эукариоты. В клетке одно или несколько ядер. | Эукариоты. Ядро чаще одно. |
| Хроматофоры (хлоропласты) разнообразны по форме, числу, строению, набору пигментов, местоположению в клетке. Лейкопласты редко. У прокариотических водорослей нет хроматофоров. | Линзовидные хлоропласты. Есть хромопласты, лейкопласты. |
| Тилакоиды в хроматофорах по одному, в ламеллах и реже в гранах (зеленые, харовые). | Тилакоиды только в гранах. |
| Разнообразные хлорофиллы: a, b, c, d, e у водорослей разных отделов. | Только a и b хлорофиллы. |
| Каротиноиды (в том числе специфические – фукоксантин, сифонеин, сифоноксантин, вашериаксантин), фикобилины (фикоцианин, аллофикоцианин, фикоэритрин). | Каротиноиды. |
| Хроматическая адаптация в водоемах. | Нет. |
| У многих есть пиреноид. Пиреноид – это белковая структура, пространственно и функционально связанная с хроматофором, представляющая собой скопление фермента фиксации углекислоты при фотосинтезе. | Пиреноид есть только у некоторых мохообразных. |
| Разнообразные запасные вещества: крахмал, хризоламинарин (лейкозин), парамилон, ламинарин, багрянковый крахмал, сахариды, многоатомные спирты, цианофициновые гранулы, цианофициновый крахмал, белки, липиды (масла, жиры), волютин, гликоген. | Часто крахмал, масло, белки. |
| Кристы митохондрий разные: пластинчатые, трубчатые, дисковидные. | Кристы митохондрий только пластинчатые (ламеллярные). |
| Центриоли есть у немногих (хлорококковые зеленые, желтозеленые, бурые). | Центриолей нет. |
| Жгутики у подвижных стадий (зооспор, гамет) разные: изоморфные, изоконтные, гетероморфные, гетероконтные. | Жгутики есть только у сперматозоидов (только изоморфные и изоконтные). |
| Размножение разнообразное: – вегетативное (деление клетки, фрагментация колонии и многоклеточного таллома, столоны, выводковые почки, гормогонии, акинеты, гонидии, кокки, планококки, клубеньки), – собственно бесполое (подвижные и неподвижные споры), – половое (хологамия, гаметогамия, конъюгация, автогамия). Изогамия, гетерогамия, оогамия истинная или примитивная. | Вегетативное (выводковые почки, вегетативные органы). Собственно бесполое только неподвижными спорами. Половое только гаметогамия – истинная оогамия, сифоногамия. Семенное размножение. |
| Спорангии одноклеточные. | Спорангии многоклеточные. |
| Споры гаплоидные и диплоидные. | Споры только гаплоидные. |
| Все споры одинаковые по размеру. | Есть одинаковые споры, но есть и разные споры – микроспоры и мегаспоры. |
| Гаметангии у большинства одноклеточные. Только у харовых исключение. | Гаметангии только многоклеточные. |
| Для копуляции гамет нужна капельно-жидкая влага. | Для копуляции гамет капельно-жидкая влага нужна только споровым. У семенных мужские гаметы доставляют гаусториальная или пыльцевая трубки. |
| Разнообразные (все из возможных) жизненные циклы и типы мейоза. Есть циклы без полового процесса и мейоза. | Только один тип цикла и мейоза – гапло-диплобионтный со спорическим (промежуточным) мейозом. |
| В гапло-диплобионтном цикле есть 1) изоморфное чередование поколений, 2) гетероморфное с преобладанием гаметофита, 3) гетероморфное с преобладанием спорофита. | Только гетероморфное чередование поколений: 1) гетероморфное с преобладанием гаметофита (только у мохообразных), 3) гетероморфное с преобладанием спорофита (у всех других высших растений). |
Уровни организации водорослей
По уровню организации водоросли делятся на одноклеточные, колониальные и многоклеточные.
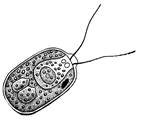
Одноклеточные водоросли – представлены одиночными клетками (талломами) и присутствуют среди всех отделов, за исключением бурых и харовых водорослей. Например, Euglena, Chlamydomonas.
Колониальные водоросли представлены в таких отделах как синезеленые, золотистые, диатомовые, желтозеленые, красные и зеленые. Колония – более или менее сложное объединение талломов, которые при делении остаются соединенными, как правило, посредством слизи, выростов покровов тела, редко при помощи плазмодесм. Например, Microcystis, Scenedesmus, Volvox, Melosira. Ценобий – это колония, в которой число талломов определяется на ранних стадиях развития и не меняется до следующей репродуктивной фазы. Талломы в составе ценобия только растут, но не делятся. Например, Scenedesmus, Volvox.
Многоклеточные водоросли имеют таллом из многих клеток. Такие водоросли есть среди многих отделов, за исключением эвгленовых, криптофитовых и диатомовых водорослей. Например, Ulothrix, Laminaria.
Морфологическая дифференциация таллома водорослей
Огромное разнообразие внешней формы водорослей может быть сведено к нескольким основным структурам, или ступеням морфологической дифференциации. Эти структуры повторяются в разных систематических группах, что указывает на некоторый параллелизм в эволюции их форм.
Монадная структура. Наиболее характерным признаком является наличие жгутиков, с помощью которых организмы активно передвигаются в воде. У простейших одноклеточных и колониальных форм монадная структура наблюдается в течение всей вегетативной жизни индивидуума, а у более высокоорганизованных бывает лишь у репродуктивных клеток. Монадный тип структуры оказался эволюционно перспективным. На его основе развились другие, более сложные структуры, связанные с утратой подвижности в вегетативном состоянии. Например, Volvox, Euglena, Chlamydomonas.
Амебоидная, или ризоподиальная структура. Наиболее важные признаки этого типа структуры – отсутствие твердой оболочки и способность к амебоидному движению с помощью псевдоподий. Амебоидный тип таллома, по мнению многих ученых, является эволюционным тупиком, возникшим в результате приспособления монадных форм к особым условиям существования в биотопах, богатых органикой. Например, Rhizochloris.
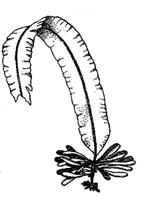
Гемимонадная (пальмеллоидная, капсальная, тетраспоральная, трансгрессивная)структура. Характерным признаком этого типа структуры является объединение неподвижных клеток в слизистую колонию. Клетки в общей слизи колонии независимы друг от друга и часто имеют органеллы, свойственные монадным организмам (сократительные вакуоли, стигмы, жгутики или их производные). Клеткам гемимонадного типа, как и монадным, свойственно полярное строение. Если эта структура встречается в жизненном цикле водоросли как временное явление, ее называют пальмеллевидным состоянием. Гемимонадный тип таллома стал важным этапом на пути морфологической эволюции водорослей в направлении от подвижных монадных к типично растительным неподвижным формам. Например, Apiocystis.
Коккоидная структура. Неподвижный за счет жгутиков и псевдоподий таллом, не изменяющий форму тела, покрытый плотной клеточной оболочкой (целлюлоза, муреин в составе стенки) или панцирем. Например, Chlorella, Microcystis, Pinnularia. Клетки различной формы (не всегда шаровидные), одиночные или соединенные в колонии. В эволюционном плане этот тип структуры рассматривается как исходный для возникновения многоклеточных талломов. У диатомовых водорослей, имеющих на панцире шов, коккоидные талломы подвижные, но за счет особого механизма, связанного с циркуляцией слизи или воды в шве.
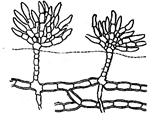
Нитчатая, или трихальная структура. Представляет талломы, состоящие из клеток, расположенных в форме нити. Нити могут быть простые (улотрикс) или разветвленные (бульбохете, эдокладиум), одно- или многорядные, свободноживущие или прикрепленные, одиночные или объединенные в разного типа соединения. Нитям присуще важнейшее свойство растительных организмов – неограниченный рост в течение вегетативной фазы жизненного цикла. Рост называют диффузным, если способностью делиться обладают все клетки нити. Деление клеток может происходит только на ограниченных участках слоевища, называемых зонами роста, или меристемальными зонами. В зависимости от положения зон роста различают интеркалярный, базальный и апикальный рост. При интеркалярном росте меристемальная зона находится в средней части нити, при базальном – у основания нити, при апикальном – рост осуществляется делением верхушечных клеток. Нижняя клетка нередко превращается в бесцветный ризоид или стопу, лишенный хлоропластов. Нитчатый тип таллома послужил отправным пунктом для развития других более сложных типов талломов. Например, Ulothrix, Spirogyra.
Разнонитчатая, или гетеротрихальная структура. Таллом данной структуры состоит из стелющейся по субстрату горизонтальной части и отходящей от нее вертикальной (трентеполия, стигеоклониум). Стелющиеся по субстрату нити выполняют главным образом функцию прикрепления, вертикальные, приподнимающиеся над субстратом нити – ассимиляционную функцию. Может наблюдаться редукция горизонтальной (Драпарнальдия) или вертикальной (Coleochaete) части таллома, реже и горизонтальной, и вертикальной частей (Desmococcus).
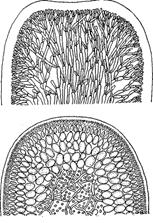
Паренхиматозная, или тканевая структура. Клетки первичной нити способны к делению в разных плоскостях. В пределах этого типа структуры можно наблюдать постепенное усложнение таллома от простых недифференцированных пластинок с диффузным ростом до сложноорганизованных слоевищ, с «тканями», выполняющими различные функции. Например, Ulva, Porphyra, Laminaria.
Псевдопаренхиматозная, или ложнотканевая структура. Характерным признаком этого типа структуры является образование достаточно крупных слоевищ в результате переплетения и срастания нитей разветвленного нитчатого (иногда сифонального) таллома. Образуются ложные ткани. Тупиковая ветвь в морфологической эволюции водорослей. Например, Batrachospermum, Nemalion.
Харофитная структура. Имеется только у харовых водорослей. Характеризуется крупными многоклеточными талломами линейно-членистого строения, которые состоят из главных побегов с ветвями («стеблей»), боковых ветвей ограниченного роста («листьев») и ризоидов. Например, Chara, Nitella.
Сифональная структура. Отличительная черта такой структуры – отсутствие внутри таллома клеточных перегородок при наличии большого числа ядер. Талломы могут быть сложно расчленены и иметь довольно крупные размеры. Например, Codium, Caulerpa, Vaucheria.
Сифонокладальная структура. Представлена многоядерными клетками, объединенными в многоклеточные талломы разной формы. Берет свое начало от сифональной структуры. Имеет место сегрегативное деление, характерной особенностью которого является разобщенность процессов кариокинеза и цитокинеза. Например, Cladophora, Valonia.
|

|

| ||||||

|

|
| ||||||
|
|
| ||||||

|

| |||||||
| Cтруктуры талломов водорослей (Л. Л. Великанов и др., 1981; C. П. Вассер и др., 1989): 1 – монадная (Сhlamydomonas sp.); 2 – амебоидная, или ризоподиальная (Rhizochloris sp.); 3 – пальмеллоидная, или капсальная (Hydrurus sp.); 4 – коккоидная (Closterium sp); 5 – нитчатая, или трихальная(Bulbochaete sp.); 6 – разнонитчатая, или гетеротрихальная (Fritschiella tuberosa); 7 – паренхиматозная, или тканевая (Alaria sp.); 8 – псевдопаренхиматозная, или ложнотканевая (Furcellaria sp.); 9 – харофитная (Chara sp.); 10 – сифональная (Botrydium sp.); 11 – сифонокладальная (Cladophora sp.) |

Let’s consider functional chart (diagram) DTS TS:
To the input of equipment comes the analog signal. This is VBC that takes bandwidth from 0,3 to 3,4 kHz.
Hibrid system serves for transition from 2-wire VBC termination to a 4-wire termination, and also it prevents VBC generation with a 2-wire termination.
The LPF-3,4 in a transmission path is used for limiting of input signal frequency band before sampling for possibility of Kotelnikov (Sampling) Theorem using.
Electronic key in a transmission path carries out sampling of signal with limited spectrum (it is PAM modulator). Working frequency (fs=8кHz) of this key (sampling frequency) is defined by Kotelnikov (Sampling) Theorem. Generation equipment (GE tr) in the transmission path closes a key on very short time (considerably less than sampling period). The given key carries out functions of the pulse-amplitude modulation (PAM).
The sampling of an initial analogue signal is carried out for TDM realization. Empty intervals between samples of one channel are filled by samples of other channels and in result a group PAM signal is formed.
On reception inverse transformation is carried out. The key in the receiving path separates samples of one channel from multichannel PAM signal.
The LPF-3,4 in the receiving path restores a continuous signal from an individual PAM signal.
|
