Полимеразная цепная реакция
Метод полимеразной цепной реакции (ПЦР), предложенный в 1983 г. Карри Муллисом (Нобелевская премия, 1993 г.), явился эпохальным
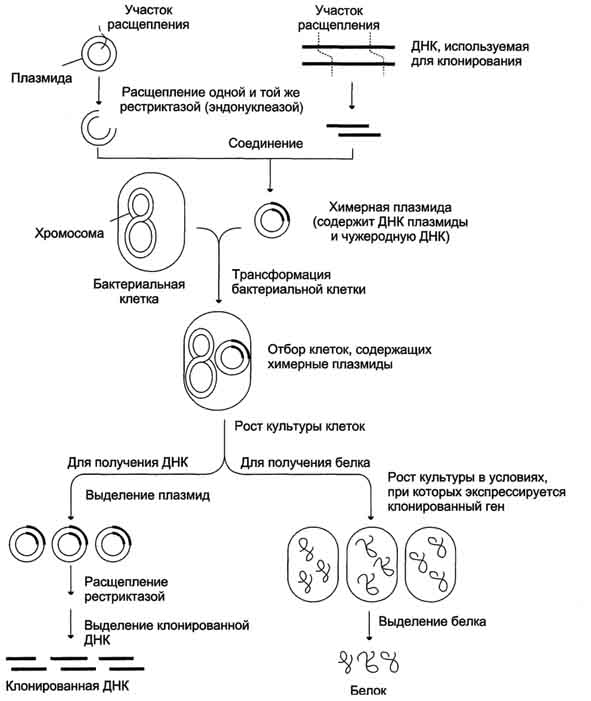
Рис. 4-66. Схема клонирования ДНК в бактериальных клетках.
открытием XX века в области молекулярной биологии. Он позволяет подвергать специфичной амплификации в условиях in vitro (в пробирке) участки ДНК длиной от нескольких десятков до нескольких сотен пар нуклеотидов, используя в качестве матрицы любые образцы ДНК. Необходимое условие для проведения ПЦР - знание нуклеотидной последовательности амплифицируемой области. Участок исследуемой ДНК гибридизуют с двумя искусственно синтезированными праймерами - олигодезоксирибонуклеотидными последовательностями длиной от 15 до 30 пар нуклеотидов, которые комплементарны 3'-концам амплифицируемого участка на кодирующей
и некодирующей нитях ДНК. Расстояние между праймерами определяет длину синтезируемых молекул. В качестве матрицы для синтеза продуктов ПЦР используют любой тип ДНК: геномную ДНК человека, различных видов про- и эукариотов, ДНК, выделенную из культур клеток, "библиотек" генов и других источников. Метод не требует больших количеств исследуемой ДНК, в принципе, достаточно даже одной молекулы, содержащейся в одном волосе на голове, одной капле крови или спермы.
Успех в разработке метода в значительной степени обусловлен использованием в качестве фермента термофильной ДНК-полимеразы, выделенной из бактерий, живущих в горячих источниках, и потому устойчивой к действию высоких температур.
Реакционная смесь для получения интересующей нас ДНК содержит исследуемую ДНК, субстраты реакции - 4 дНТФ, 2 праймера, термостабильную, или Taq-полимеразу и буфер, содержащий ионы Mg2+.
Один цикл полимеризации включает 3 этапа (рис. 4-67):
· плавление:на этой стадии реакционную смесь нагревают до температуры 90-97 °С. Исследуемая двуцепочечная ДНК денатурирует и переходит в однонитевую форму;
· гибридизация или отжиг ДНК с праймерами.В результате снижения температуры до 50-60 °С происходит комплементарное связывание праймеров с цепями матричной ДНК и образование двухцепочечного участка на каждой из нитей ДНК;
· элонгация,удлинение нитей ДНК, комплементарных матричной ДНК, катализирует Taq-полимераза в направлении от 5'- к 3'-концу.
Затем снова наступает этап плавления, когда за счёт повышения температуры синтез ДНК прекращается, и двунитевой участок между матричными и вновь синтезированными молекулами ДНК денатурирует. Во втором и последующих циклах праймеры гибридизируются с исходной матричной ДНК и с вновь синтезированными молекулами ДНК, количество которых нарастает в геометрической прогрессии. В последнем случае синтез ДНК заканчивается не из-за изменения температурного режима, а по достижении ДНК-полиМеразой границы амплифицированного участка, что определяет строго определённый размер продукта с точностью до одного нуклеотида.
Каждый из этапов цикла имеет продолжительность от десятков секунд до 1-3 мин, в результате полный цикл длится от одной до нескольких минут.
Описанную процедуру амплификации ДНК проводят в автоматическом режиме в приборе - циклизаторе, или термоциклере, амплификаторе ДНК. Такой прибор позволяет задавать нужное количество циклов и выбирать оптимальные временные и температурные параметры. За 25-30 циклов число синтезированных копий ДНК достигает нескольких миллионов.
С помощью ПЦР можно получить достаточное количество копий участков ДНК, в которых предполагаются присутствие мутаций, полиморфизм сайтов, можно проводить ДНК-диагностику инфицированности пациентов вирусными, бактериальными и грибковыми возбудителями болезней.
Д. ДНК-диагностика заболеваний
Используя технику рекомбинантных ДНК, удаётся исследовать варианты генов, ответственных за развитие многих заболеваний. Этим способом идентифицированы точечные мутации, вызванные заменой одного азотистого основания, делениями или вставками, приводящими к появлению аллелей, кодирующих функционально неактивные белки. Дефектные "полиморфы" возникают как за счёт изменений в кодирующих участках гена, так и в результате мутаций в некодирующих; областях, тесно примыкающих к генам и вызьшающих нарушение их работы.
Разработанные технологии позволяют вести целенаправленное картирование генов человека в рамках международного проекта "Геном человека". Официально эта научная программа с участием ведущих молекулярно-генетических лабораторий США, стран Западной Европы, а также России и Японии оформилась в 1990 г. В ходе работы над проектом картированы 923 гена, вызывающих развитие моногенных заболеваний, более 100 из них полностью секве-нированы. К концу 2001 г. работами лабораторий США, Великобритании, Японии и ряда европейских стран с точностью до 90% завершена расшифровка генома. Ожидается, что в течение ближайших 2-3 лет будут изучены все гены, ответственные за развитие патологических

Рис. 4-67. Полимеразная цепная реакция.
процессов у человека. Это позволит вывести диагностику и лечение многих болезней на новый уровень.
Остановимся на некоторых методах, широко используемых для идентификации моногенных болезней.
1. Полиморфизм длины рестрикционных
фрагментов (ПДРФ)
Мутации, возникающие в участках узнавания определённых рестриктаз, делают эти участки ДНК нечувствительными к действию ферментов. Это может быть легко обнаружено по изменению длины рестрикционных фрагментов ДНК. ПДРФ-анализ включает следующие этапы: выделение геномной ДНК, её рестрикцию специфической эндонуклеазой, электрофоретическое разделение образующихся фрагментов ДНК и идентификацию этих фрагментов путём блот-гибридизации по Саузерну. На электрофореграммах при отсутствии рестрикции в исследуемой ДНК выявляют один крупный фрагмент, соответствующий по длине последовательности ДНК между двумя соседними участками рестрикции для той же эндонуклеазы. При наличии рестрикции в полиморфном участке на электрофореграмме будет присутствовать меньший по размерам фрагмент, равный расстоянию между полиморфным участком рестрикции и одним из ближайших постоянных участков рестрикции (рис. 4-68).
ПДРФ-анализ может быть значительно упрощён в том случае, если возможна специфическая амплификация участка ДНК, содержащего полиморфный сайт рестрикции. Тестирование состояния этого локуса возможно путём проведения ПЦР и рестрикции амгошфицированного фрагмента. При отсутствии сайта узнавания в исследуемой области ДНК размеры амплифицированного фрагмента не изменятся после его обработки рестриктазой. Если участок узнавания не изменён, обработка ферментом приведёт к образованию 2 маленьких фрагментов с той же суммарной длиной, что и исходный фрагмент.
При обследовании пациентов и членов их семей на носительство патологических генов широко используют этот метод, с помощью которого:
- идентифицируют делеции в гене дистрофина, на долю которых приходится около 60% всех мутаций, вызывающих миодистрофию Дюшенна;
- диагностируют гемофилию А, некоторые та-лассемии, ретинобластому и гранулематоз;
- контролируют здоровье детей в семьях, в которых родители являются гетерозиготами по гену серповидно-клеточной анемии и другим дефектным генам.
2. Определение мутаций с помощью
аллельспецифических проб
Олигонуклеотиды, аллельспецифичные по определённым мутациям, можно использовать в качестве праймеров в ПЦР при клиническом тестировании населения на наличие патологического гена. Если ДНК, полученная от пациента, амплифицирует с мутантным олигонулеотидом, то следовательно пациент является носителем мутации. Если нуклеотидная последовательность в исследуемом гене не изменена, то олигонуклеотид, содержащий мутацию, не свяжется с ДНК-матрицей, и ПЦР не пойдёт.

Рис. 4-68. Рестрикционный анализ ДНК гемоглобина человека, больного серповидно-клеточной анемией (HbS). Мис-сенс-мутация, ответственная за возникновение серповидно-клеточной анемии, связана с заменой в гене β-цепи глобина триплета GAG на GTG. При этом утрачивается участок рестрикции фермента Mstll, узнающего последовательность CCTNAGG, где N может быть любым основанием. При наличии мутации генный зонд гибридизуется с более крупным фрагментом ДНК размером 1,3 килобазы, имеющим при электрофорезе меньшую подвижность, чем продукт рестрикции нормального гена, длина которого равна 1,1 килобазы.
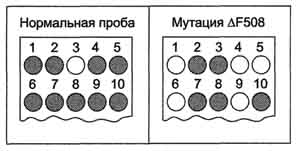
Рис. 4-69. Генное зондирование на носительство муко-висцидоза. С помощью ПЦР амплифицируют геномную ДНК, и продукт переносят на 2 нейлоновых фильтра, один из которых гибридизуют с P32Р-зондом на нормальный аллель, а второй с 32Р-зондом на мутантный аллель ΔF508. Образование гибридов обнаруживают радиоавтографически. Пробы 1-3 служили контролями: первая содержала продукты ПЦР ДНК от гомозигот по нормальному гену; вторая - от гетерозиготного носителя мутации ΔF508; третья - от больного муковисцидозом, гомозиготного по мутации ΔF 508. Пробы 4-10 получены при обследовании 7 пациентов на носительство ΔF 508: 4, 5, 6 и 9 оказались гомозиготами по нормальному гену, а 7, 8 и 10 - гетерозиготными носителями мутантного гена.
Е. Использование ДНК-технологий для получения лекарственных препаратов и лечения различных болезней
Вакцины- очищенные белки, антигенные детерминанты ряда возбудителей вирусных и бактериальных инфекций. В последнее время их получают, пользуясь техникой рекомбинантных ДНК. Первой вакциной, синтезированной этим способом, была вакцина против вируса гепатита В.
Белки, имеющие терапевтическое значение,получают с использованием этой технологии во многих странах мира. Так, одним из первых синтезирован инсулин человека (рис. 4-70). В клетках Е. coli, трансформированных гогазмидами, содержащими ДНК, которая кодировала А- и В-цепи инсулина, нарабатывают белковые продукты А- и В-цепей. После очистки их подвергают фолдингу и окислению, которое обеспечивает образование соответствующих дисульфидных мостиков.

Рис. 4-70. Получение инсулина человека в клеткахЕ. coli. 1 - трансформация клеток Е. coli плазмидами, которые содержат гены, кодирующие структуру А- и В-цепей инсулина; 2 - синтез А- и В-цепей инсулина в процессе выращивания культуры трансформированных клеток Е. coli; 3 - выделение и очистка А- и В-цепей инсулина; 4 - пространственная укладка А- и В-цепей (лнсулина и окисление остатков цистеина.
Аналогичным способом получен гормон роста, используемый для лечения детей с недостаточностью этого гормона. Более сложные белки получены в культуре клеток млекопитающих. Так, дефекты в гене фактора VIII, кодирующего один из белков - участников свёртывающей системы крови, ответственны за возникновение гемофилии. До того как фактор VIII был получен методами генной инженерии, большое количество больных погибало от СПИДа или гепатита, которыми они заражались в результате введения выделенного из крови фактора
VIII или переливания крови от доноров, являвшихся носителями этих болезней.
Тканевый активатор плазминогена (ТАЛ) -протеаза, участвующая в процессе фибриноли-за и предотвращающая образование тромбов в кровеносном русле; получена с помощью рекомбинантных ДНК. ТАП назначают больным с ишемической болезнью сердца для ускорения растворения тромбов, которые могут вызвать закупорку коронарных артерий и нарушить поступление кислорода в миокард.
Осуществлено получение рекомбинантных факторов роста, обеспечивающих восстановление гемостаза: эритропоэтина, интерлейкинов, колоний-стимулирующих факторов. Эти препараты используют в лечении больных анемией, после трансплантации костного мозга или химиотерапии, чтобы стимулировать образование клеток крови и снизить риск иммунодефицита. Разработаны методы получения белков человека с использованием трансгенных животных; эти белки получают в результате искусственного введения чужеродного гена в оплодотворённую яйцеклетку или в ранние зародыши млекопитающих (рис. 4-71). Генноинженерные мероприятия можно провести таким образом, чтобы интересующий нас белок человека секретиро-вался с белками молока.
Генная терапия- лечение наследственных, многофакторных и инфекционных заболеваний путём введения в соматические клетки пациентов генов, которые обеспечивают исправление генных дефектов или придают клеткам новые функции.
Первый клинический опыт применения генной терапии был осуществлён в 1990 г. в Бетесде (США) на четырёхлетней девочке, страдавшей наследственным иммунодефицитом, вызванным мутацией в гене аденозиндезаминазы (ADA.) Ребёнку были введены её собственные лимфоциты, предварительно трансформированные вне организма генной конструкцией, включающей ген ADA + ген пео + ретровирусный вектор. Лечебный эффект наблюдался в течение нескольких месяцев, после чего процедуру введения гена повторяли многократно без видимых неблагоприятных эффектов.
Для успешной генотерапии необходимо:
- обеспечить эффективную доставку чужеродного гена в клетки-мишени;
- создать условия для длительной экспрессии гена в этих клетках.
К настоящему времени разработаны химические, физические и биологические методы доставки чужеродного гена в клетки-мишени. Однако пока только вирусные векторы или генетические конструкции, включающие вирусные последовательности, способны к эффективной доставке необходимого гена и его последующей длительной экспрессии. В результате из более чем 175 уже одобренных протоколов клинических испытаний по генотерапии более 120 основаны на применении ретровирусных векторов.
В геном пациента чужеродная ДНК может вводиться либо в культуре клеток (ex vivo), либо непосредственно в организм больного (in vivo). При осуществлении первого способа выделяют и культивируют специфический тип клеток пациента, вводят в него чужеродный ген, отбирают трансформированные клетки и реинфузируют их тому же больному (рис. 4-72).
Генная терапия in vivo основана на прямом введении в специализированные ткани больного клонированных и определённым образом упакованных последовательностей ДНК, поступающих с помощью рецепторов в определённые типы клеток. В этом способе гены вводят, как правило, в виде аэрозольных и инъецируемых форм. Наиболее часто аэрозольную генотерапию используют при лечении болезней лёгких (например, раке лёгких) и муковисцидоза.
Наряду с развитием исследований, касающихся лечения наследственных дефектов, генотерапию всё чаще используют для лечения ненаследственных, главным образом, инфекционных и онкологических болезней (см. раздел 16).
Единственное и непременное ограничение таких работ состоит в том, чтобы все генотерапевтические мероприятия были направлены на конкретного больного и затрагивали только его соматические клетки.
Современный уровень знаний не позволяет проводить коррекцию генных дефектов на уровне половых клеток и клеток ранних доимплантационных зародышей человека в связи с реальной опасностью засорения генофонда нежелательными генными конструкциями и внесения мутаций с непредсказуемыми результатами.
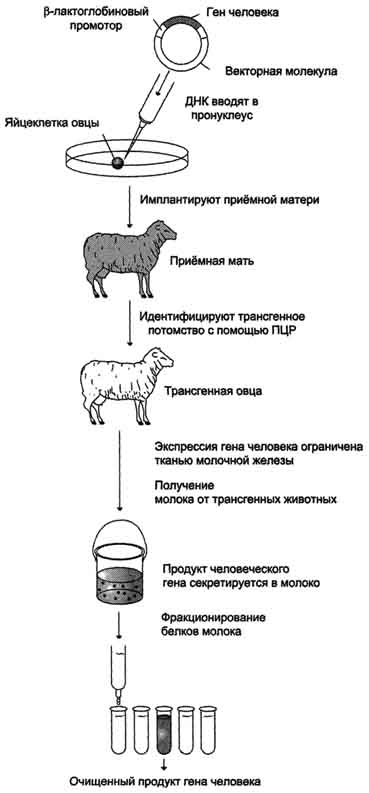
Рис. 4-71. Использование трансгенных животных для получения белков человека. Ген человека встраивают в вектор таким образом, чтобы он был под контролем β-лактоглобинового промотора, который активен только в клетках молочной железы. Присутствие у трансгенного потомства гена человека контролировали с помощью метода ПЦР, в которой использовали праймеры к гену человека. При фракционировании белков молока получают белковый продукт экспрессии гена человека.
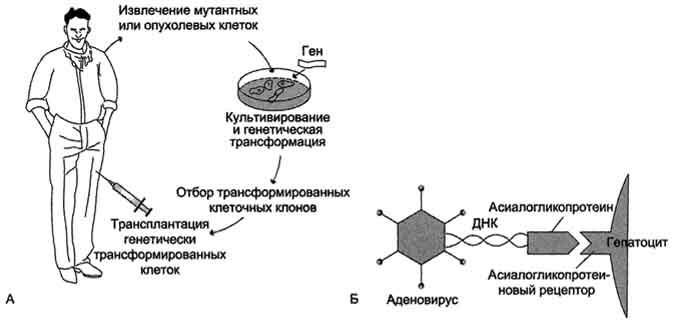
Рис. 4-72. Введение чужеродного генаex vivo(А) иin vivo(Б). А. Введение чужеродного "лечебного" гена в организм больного в составе клеток, содержащих этот ген. Б. Введение "лечебного" гена в составе конструкции, содержащей: ДНК, включающую этот ген; белок (например асиалогликопротеин), взаимодействующий с соответствующим рецептором на мембране клеток; вирусный вектор (аденовирус), обеспечивающий длительную экспрессию "лечебного" гена.
В целях предотвращения распространения дефектных генов в популяции людей и рождения детей с наследственными патологиями во многих странах мира работают генетические консультанты, а также проводят пренатальную диагностику, позволяющую оценить здоровье плода с использованием анализа ДНК на самых ранних стадиях развития.
