Экзонуклеазная активность ДНК-пол.
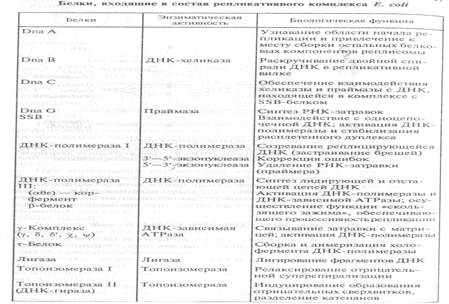
ДНК-полимеразы присутствуют во всех клетках про- и эукариот. Некоторые прокариотические и эукариотические ДНК-полимеразы выделены в чистом виде, и их ферментативные и физические свойства охарактеризованы. Физические свойства этих ферментов не совсем идентичны, но механизм катализа, который они существляют, в общих чертах одинаков: каждая из них способна удлинять цепь ДНК, наращивая ее посредством присоединения каждого последующего нуклеотида к 3’-концу цепи ДНК или РНК.Основные белки и ферменты, входящие в состав
репликативного комплекса E. сoli, а также их функции, указаны в таблице. У E. сoli есть три ДНК-полимеразы– I, II и III (названия даны по мере их открытия).
ДНК-пoлимераза I E. coli
Выделена А. Корнбергом и сотр. в 1958 г. Изучена наиболее полно. Представляет собой
одиночный полипептид с мультифункциональными активностями. Для осуществления
реакции полимеризацииферменту необходим праймер, содержащий свободную 3’-ОН-
группу и матрица, детерминирующая присоединение нужного нуклеотида.
Экзонуклеазные реакции ДНК-полимеразы I. 3’→ 5’-экзонуклеазная активность
Кроме полимеризации ДНК-пол.I катализирует две другие реакции. В ходе одной из них происходит гидролиз фосфодиэфирных связей в одной из цепей ДНК, начиная с 3’-конца цепи (3’→ 5’-экзонуклеазная активность), причем, за один акт удаляется один нуклеотид, начиная с 3’-конца цепи. Рисунок
Втораяреакция также заключается в отщеплении нуклеотидов, но
гидролиз идет с 5’-конца цепи ДНК к 3’-концу (5’→ 3’-экзонуклеазная активность).
Различные активности ДНК-пол.I принадлежат разным участкам
полипептидной цепи с молекулярной массой 109 000 дальтон.
Большой С-концевой участок(76 000 дальтон) проявляет 5’→3’-
полимеразную и 3’→ 5’-экзонуклеазную активности.
Малый, N-концевой, фрагмент (36 000 дальтон) обладает только
5’→ 3’-экзонуклеазной активностью.
Большой фрагмент называется также фрагментом Кленова, он способен инициировать репликацию in vitro. 3’→ 5’-экзо-нуклеазная активность обеспечивает контроль за присоединением каждого последующего нуклеотида и удаление ошибочно вставленного нуклеотида с растущего конца цепи ДНК. С помощью 5’→ 3’-экзонуклеазной активности вырезаются праймеры.
Две другие ДНК-полимеразы присутствуют в клетках E. сoli в меньших количествах.
ДНК-пол. II (мол.масса 90 кДа) представлена одной полипептидной цепью, обладает полимеразной и 3’→ 5’-экзонуклеазной активностями. Она плохо соединяется с одноцепочечными ДНК, но лучше работает с биспиральной ДНК, имеющей одноцепочечные бреши длиной в несколько десятков нуклеотидов, обладает лишь 10 %-й ДНК-полимеразной активностью по сравнению с ДНК-полимеразой I. Предполагают, что основной функцией ДНК-полимеразы II является достраивание поврежденных участков в молекуле ДНК, т. е. репарация ДНК. Она может заполнять пробелы между фрагментами ДНК за счет полимеразной активности, но не способна отщеплять РНК-нуклеотиды от фрагментов Оказаки (т.к. не обладает 5’→3’-экзонуклеазной активностью) или осуществлять ник-трансляцию.