График проведения обзорных лекций 6 страница
В конечном счете экосистему можно рассматривать как сумму популяций (или их частей) разных видов, взаимосвязанных между собой и находящихся в тесном единстве с окружающей средой.
Несмотря на абстрактность вычленения популяций из общих систем (биоценозов, экосистем), им свойственен ряд специфических закономерностей функционирования, важных для существования вида и в целом экосистем различного ранга. На уровне популяций происходят основные адаптации, естественный отбор и эволюционные процессы. Разнообразие популяций внутри вида резко увеличивает его приспособительные способности, освоение среды и, в конечном счете, возможности выживания.
Проявляя заботу о сохранении вида, человек должен прежде всего думать о сохранении популяций. Для популяций различных видов существуют допустимые пределы снижения численности особей, за которыми существование популяции становится невозможным. Точных данных о критических значениях численности популяций в литературе нет. Приводимые значения разноречивы. Остается, однако, несомненным факт, что чем мельче особи, тем выше критические значения их численности. Для микроорганизмов это миллионы особей, для насекомых - десятки и сотни тысяч, а для крупных млекопитающих - несколько десятков. Численность не должна уменьшаться ниже тех пределов, за которыми резко снижается вероятность встречи половых партнеров. Критическая численность также зависит от других факторов. Например, для некоторых организмов специфичен групповой образ жизни (колонии, стаи, стада). Группы внутри популяции относительно обособлены. Могут иметь место такие случаи, когда численность популяции в целом еще достаточно велика, а численность отдельных групп уменьшена ниже критических пределов. Например, колония (группа) перуанского баклана должна иметь численность не менее 10 тыс. особей, а стадо северных оленей - 300-400 голов.
V.1. Структура популяций
Для понимания механизмов функционирования и решения вопросов использования популяций важное значение имеют сведения об их структуре.
Различают половую, возрастную, территориальную и другие виды структуры. В теоретическом и прикладном планах наиболее важны данные о возрастной структуре, под которой понимают соотношение особей (часто объединенных в группы) различных возрастов.
Обычно наибольшей жизнеспособностью отличаются популяции, в которых все возраста представлены относительно равномерно. Такие популяции называют нормальными. Если в популяции преобладают старческие особи, это однозначно свидетельствует о наличии отрицательных факторов в ее существовании, нарушающих воспроизводительные функции. Такие популяции рассматривают как регрессивные, или вымирающие. Требуются срочные меры по выявлению причин такого состояния и их исключению. Популяции, представленные в основном молодыми особями, рассматриваются как внедряющиеся, или инвазионные. Жизненность их обычно не вызывает опасений, но велика вероятность вспышек чрезмерно высокой численности особей, поскольку в таких популяциях не сформировались трофические и другие связи. Особенно опасно, если такие популяции представлены видами, которые здесь ранее отсутствовали. В таком случае популяции обычно находят и занимают свободную экологическую нишу и реализуют свой потенциал размножения, интенсивно увеличивая численность.
Если популяция находится в нормальном или близком к нормальному состоянии, человек из нее может изымать то количество особей или биомассу (последний показатель обычно используется применительно к растительным сообществам), которая прирастает за промежуток времени между изъятиями. Ясно, что изыматься должны прежде всего особи послепродуктивного возраста (окончившие размножение). Если преследуется цель получения определенного продукта, то поправки на возраст, пол или другие характеристики популяций корректируются с учетом поставленной задачи.
Эксплуатация популяций растительных сообществ, в частности, с целью получения древесного сырья обычно приурочивается к тому периоду, когда имеет место возрастное замедление накопления продукции (прироста). Этот период обычно совпадает с максимальным накоплением древесной массы на единице площади.
Количество изымаемой продукции и способ ее изъятия сообразуется с биологическими особенностями популяций. Например, у животных, ведущих групповой образ жизни, как отмечалось выше, нельзя уменьшать численность групп до такой степени, которая повлекла бы за собой потерю ими свойств оптимизации жизненных процессов.
Лесоводами применительно к решаемым задачам и сообразуясь с эколого-биологическими свойствами популяций (экосистем) разработаны различные виды рубок. Прежде всего, они делятся на две большие группы: промежуточного и главного пользования. Рубки промежуточного пользования проводятся практически во всех возрастах жизни леса. При их проведении, наряду с изъятием продукции, преследуется цель создания более благоприятных условий для жизнедеятельности и роста остающейся части древостоя. Ими же создаются условия для увеличения доли более ценных с точки зрения целей хозяйства видов (например, хвойных в смешанных хвойно-лиственных сообществах).
При рубках главного пользования убирается весь древостой, достигший возраста спелости. Этот урожай может сниматься единовременно (сплошные рубки) или в несколько приемов (постепенные, выборочные рубки). Изымается при этом такая часть древостоя, которая не нарушила бы жизнедеятельности популяций и экосистем в целом, механизмов их самоподдержания и соморегулирования (гомеостаза). Такой тип ведения хозяйства рассматривают как мягкое управление природными процессами.
С целью создания условий для появления молодого поколения леса при сплошных рубках применяются такие лесоводственные приемы, как вырубка последовательно небольшими площадями (лесосеками). В таком случае остающиеся рядом с вырубкой древостой являются источником семян, умеренного притенения появляющегося молодого поколения леса, препятствием буйному росту конкурентов из трав, кустарников и нежелательных древесных растений. Появлению молодого поколения леса способствует также оставление на вырубках отдельных, как правило, лучших деревьев, которые выполняют роль обсеменителей и носят название семенников.
Однако в обширных лесных массивах Севера и других регионов часто проводятся так называемые концентрированные рубки большими площадями без учета возможностей восстановления их молодым поколением леса. Они проводятся с использованием тяжелой техники, сопровождаются сильным разрушением и уплотнением почвенного покрова. Это, в свою очередь, ведет, как правило, к цепным реакциям природных процессов, в частности, сложившиеся круговороты воды сменяются накоплением застойных вод на поверхности почв с последующей сменой лесных экосистем болотными. В других случаях, например на песчаных почвах, следствием подобного вмешательства в экосистемы является опустынивание или полное разрушение экосистемы. Такой тип ведения хозяйства рассматривается как жесткое вмешательство в природные процессы. Оно не должно иметь места в деятельности человека.
V.2. Динамика популяций. Гомеостаз
К числу важнейших свойств популяций относится динамика свойственной им численности особей и механизмы ее регулирования. Всякое значительное отклонение численности особей в популяциях от оптимальной связано с отрицательными последствиями для ее существования. В связи с этим популяции обычно имеют адаптационные механизмы, способствующие как снижению численности, если она значительно превышает оптимальную, так и ее восстановлению, если она уменьшается ниже оптимальных значений.
Каждой популяции и виду в целом свойственен так называемый биотический потенциал, под которым понимают теоретически возможное потомство от одной пары особей при реализации способности организмов к биологически обусловленному размножению. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так, дрожжевые клетки, размножающиеся простым делением, при наличии условий для реализации биотического потенциала могли бы освоить все пространство земного шара за несколько часов; гриб дождевик, приносящий до 7,5 млрд. спор, уже во втором поколении освоил бы весь земной шар. Крупным организмам с низким потенциалом размножения потребовалось бы для этого несколько десятилетий или столетий.
Однако биотический потенциал реализуется организмами со значительной степенью полноты только в отдельных случаях и в течение коротких промежутков времени. Например, если быстроразмножающиеся организмы (насекомые, микроорганизмы) осваивают какой-либо субстрат или среду, где нет конкурентов. Такие условия создаются, в частности, при освоении экскрементов крупных животных насекомыми, при размножении организмов в средах, богатых питательными веществами, например в загрязненных органическими или биогенными веществами водоемах и т. п. В этом случае увеличение численности идет по j-образной кривой. Такой тип роста носит название экспоненциального. Близкий к экспоненциальному тип роста характерен в настоящее время для популяции человека. Он обусловлен прежде всего резким снижением смертности в детском возрасте. Для человека характерна кривая увеличения численности первого типа (рис. 6).
Рис. 6
Экспоненциальная (А) и логистическая (Б) кривые роста популяций
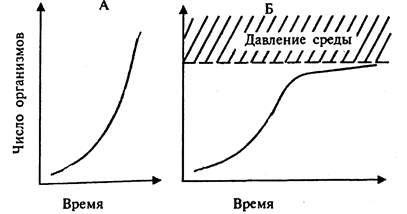
Для большинства же популяций и видов рост численности характеризуется кривой второго типа, которая отражает высокую смертность молодых особей или их зачатков (яйца, икринки, споры, семена и т. п.). При таком типе выживаемости (смертности) численность популяции обычно выражается s-образнои кривой (см. рис.6). Такая кривая носит название логистической. Но и в этом случае периодические колебания численности особей значительны. Такие отклонения от средней численности имеют сезонный (как у многих насекомых), взрывной (как у некоторых грызунов - лемминги, белки) или сглаженный (как у крупных млекопитающих) характер. Численность при этом может существенно отличаться от средних значений: для насекомых - в 107- 106 раз, для позвоночных животных, например грызунов, - в 105 - 106 раз.
Периоды резкого изменения численности носят название «популяционных волн», «волн жизни», «волн численности». Причины таких колебаний до конца не ясны. В одних случаях их связывают с пищевым фактором, в других - с погодными (климатическими) явлениями (например, для леммингов - с количеством тепла, приносимого Гольфстримом), в третьих - с солнечной активностью или комплексом взаимосвязанных факторов, что наиболее вероятно.
Резкие изменения численности относительно средних значений имеют обычно отрицательные следствия для жизни популяций: при высокой численности - из-за ослабления всех особей в результате недостатка пищи, самоотравления среды, возможных массовых заболеваний и т. п.; при низкой численности - из-за превышения порога ее минимальных значений.
В целом можно выделить динамику популяций, независимую от плотности (численности) ее особей и зависимую от плотности. Для первого типа характерна отмеченная выше экспоненциальная кривая роста. Для второго - логистическая кривая. Соответственно различаются и механизмы (факторы), оказывающие влияние на плотность (численность) особей. При независимом от плотности типе изменения численности последняя обусловливается в основном абиотическими факторами (погодные явления, наличие пищи, различного рода катастрофы и т. п.). Эти факторы могут обеспечивать условия как для неограниченного, хотя и кратковременного роста популяций, так и для снижения их численности до нулевой. Такие факторы обычно называют модифицирующими (лат. модификацио-изменение).
Зависимая от плотности динамика популяций обеспечивается биотическими факторами. Их называют регулирующими. Они «работают» по принципу обратной отрицательной связи: чем значительнее численность. Тем сильнее срабатывают механизмы, обусловливающие ее снижение, и наоборот - при низкой численности сила этих механизмов ослабевает и создаются условия для более полной реализации биотического потенциала. Факторы такого типа лежат в основе популяционного гомеостаза, обеспечивающего поддержание численности в определенных границах значений.
К числу регулирующих факторов относится, в частности, взаимоотношение организмов типа хищник-жертва. Высокая численность жертвы создает условия (пищевые) для размножения хищника. Последний, в свою очередь, увеличив численность, снижает количество жертвы. Численность обоих видов в результате этого носит синхронно-колебательный характер. Регулирующие факторы, в отличие от модифицирующих, никогда не доводят численность популяций до нулевых значений вследствие того, что сила их действия уменьшается по мере уменьшения численности популяций.
Вообще действие регулирующих факторов можно рассматривать на уровне межвидовых и внутривидовых (внутрипопуляционных) взаимоотношений организмов.
К межвидовым механизмам гомеостаза относятся отмеченные выше взаимоотношения типа хищник-жертва. В таком же плане действуют и взаимоотношения паразит- хозяин. При высокой численности создаются условия для увеличения количества паразитов и паразитарных заболеваний как в результате скученности, так и вследствие ослабления организмов. К межвидовым механизмам относится также конкуренция, острота которой находится в прямой зависимости от численности организмов.
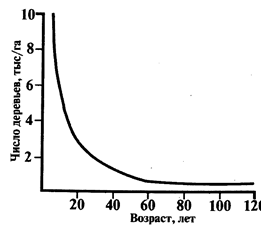
Конкуренция лежит и в основе внутрипопуляционного гомеостаза. Она здесь может проявляться в жестких и смягченных формах. Жесткие формы обычно заканчиваются гибелью части особей. В растительном мире это проявляется в явлениях так называемого самоизреживания фитоценозов. Например, на стадии всходов и молодых растений в лесных сообществах на одном гектаре насчитывается до нескольких сотен тысяч древесных растений. К возрасту спелости (100-120 лет для хвойных видов и 50-70 лет для лиственных) число экземпляров обычно не превышает 1000 на 1 га, но чаще исчисляется несколькими сотнями. Остальные погибают в процессе острой конкурентной борьбы (рис. 7). В результате этого, с одной стороны, освобождается пространство для остающихся более сильных особей, а с другой - ослабленные и погибающие особи выполняют положительную роль для сохранения популяции через включение в процессы круговорота, обогащение почвы минеральными веществами и гумусом. Часть ослабленных особей еще при жизни становится донорами для питания более сильных экземпляров. Это возможно в результате имеющего место срастания корней. Частным подтверждением таких явлений служат не единичные случаи нарастания годичных колец на пнях деревьев («растущие пни»).
Рис. 7
Ход самоизреживания соснового леса с возрастом (по Г. Ф. Морозову).
В животном мире результат острой внутривидовой борьбы проявляется часто в форме каннибализма (поедания себе подобных). Такие явления наиболее часты среди хищников. Например, взрослые окуни при высокой численности популяции, особенно в небольших водоемах, начинают питаться мальками своего вида. Явления каннибализма характерны также для некоторых грызунов, личинок насекомых, особенно в случаях существования в ограниченном пространстве. Поедание потомства домашними животными, по-видимому, один из случаев атавизма (лат. атавус - отдаленный предок) данного явления, которое раньше имело место в природных популяциях.
Смягченные формы внутривидовой конкуренции проявляются обычно через ослабление части особей, выключения их из процессов размножения. Случаи гибели особей при таких формах борьбы менее вероятны. К таким механизмам внутрипопуляционного гомеостаза относятся угнетающие (ингибирующие) выделения веществ во внешнюю среду более сильными особями, стрессовые явления, разграничение территорий (территориальность), миграции между популяциями.
Выделения во внешнюю среду характерны как для растительных, так и для животных организмов. Показано, в частности, что молодое поколение леса не появляется под материнским пологом либо находится в сильно угнетенном состоянии не только в результате дефицита светового и других факторов (минеральное питание, влагообеспеченность и т. п.). Опытным путем было установлено, что в этих процессах существенную роль играют ингибирующие выделения корней, а в ряде случаев атмосферные осадки, обогащенные химическими элементами и соединениями, вымывающимися из надземных органов взрослых растений. Препятствием для молодого поколения может также являться мощный слой слаборазложившегося мертвого органического вещества (лесной подстилки), препятствующий прорастанию семян и укоренению всходов. В спелых лесах сила влияния этих факторов обычно уменьшается, и молодое поколение постепенно сменяет теряющих конкурентную способность особей.
На примере лабораторных животных (крысах, мышах) показано, что воздух, подаваемый из помещений, где имеет место перенаселенность, в помещения, где животные свободно размещаются и нормально развиваются, приводит к замедлению роста и угнетению последних. Аналогичные результаты наблюдались в опытах с головастиками лягушек, когда в аквариумы, где они хорошо развивались, добавляли воду из аквариумов, в которых плотность организмов была чрезвычайно высокой.
Явления территориальности наиболее четко выражены в животном мире. Сюда относятся различные способы охраны занимаемых территорий. Например, пение птиц - это прежде всего сигнал о занятости территории в период размножения и последующего выкармливания потомства. У кошачьих и собачьих территориальность проявляется через мечение границ участков выделением желез, мочой или механическими отметинами на деревьях, почве и т. п.
При высокой скученности особей в популяциях регулирующим механизмом численности могут являться стрессовые явления. Они наиболее характерны для млекопитающих. При стрессах обычно часть особей снижает или теряет репродукционные функции (выключается из процессов размножения). Более сильные особи в меньшей степени подвержены стрессу и его следствиям. При ослаблении или прекращении стрессовых явлений организмы обычно восстанавливают функции жизнедеятельности и репродукционного процесса.
Миграции как фактор гомеостаза проявляются чаще всего в двух видах. Первый из них относится к массовому исходу особей из популяции при явлениях перенаселенности (нашествия). Такие явления особенно характерны для леммингов, белок и некоторых других видов с взрывным типом динамики численности. Особи, оставившие популяцию (как правило, молодняк), обычно не возвращаются на прежнее место. Значительное количество их погибает при подобных перемещениях.
Второй вид миграций связан с более постепенным (спокойным) уходом части особей в другие популяции с меньшей плотностью населения.
Вообще и при численности, близкой к оптимальной, популяции обмениваются, хотя и в меньшей степени, особями. Это важно как для исключения или уменьшения вероятности близкородственного скрещивания, так и для обмена информацией, которая имеет свою специфику в разных популяциях.
Неоднозначны реакции популяций на иммигрантов. В периоды высокой численности они препятствуют вселению особей из других популяций. При низкой численности имеет место явление противоположного порядка: резко уменьшается количество особей, оставляющих популяции, снимаются механизмы, препятствующие вселению особей из других популяций (иммигрантов).
Среди насекомых выселительная способность связана часто с появлением специфических особей, выделяемых обычно в отдельную фазу, характеризующихся большей подвижностью, стремлением к перемещениям. У тлей, например, появляется фаза с хорошо развитыми крыльями, у пустынной саранчи, кроме лучшего развития летательного аппарата, подвижность увеличивается за счет более темной окраски и в связи с этим лучшей прогреваемости тела, что для холоднокровных организмов является важнейшим фактором усиления активности.
Ясно, что гомеостаз в полной мере проявляется, если срабатывают все механизмы, лежащие в его основе. Например, не нарушается резко соотношение численностей хищников и жертв, не имеет места действие факторов, ослабляющих популяции (загрязнение, нарушение местообитаний и др.), не превышаются критические пределы численности и т. п.
В настоящее время подобные нарушения гомеостатических механизмов вызываются в большинстве случаев антропогенными факторами. В связи с этим одной из важнейших задач человека является исключение или резкое снижение действия подобных факторов. Решение вопросов такого плана относится прежде всего к области прикладной экологии, знакомству с которой посвящен следующий раздел настоящего пособия.
Пульпа зуба: вариант рыхлой соединительной ткани. Особенности фибробластов пульпы. Спектр клеток защиты (макрофаги, антиген-представляющие клетки, лимфоциты, тучные клетки).
Пульпа – разновидность рыхлой соединительной ткани, сопровождающая сосуды и нервы. Похожа на эмбриональную. Она слегка уплотняется по направлению к корневой части зуба. Плотная часть пульпы более устойчива к микроорганизмам.
В составе пульпы обнаружено 5% неорганических веществ, 40% органических и ~60% воды. Пульпа богата коллагеном (около 24,5 %), содержит сиаловую кислоту (около 0,06%), холестерин, ФЛ, особенно ФЛ кислого характера. Кислые ФЛ, в том числе фосфатидилсерин, влияют на транспорт ионов Са2+, образуя с ними комплексы, которые, в свою очередь, могут связываться с белками. Считают, что существует комплекс «белок-сахар-липид-кальций», участвующий в процессах кальцификации.
Функции пульпы: трофическая, а также участие в защитной и репаративной функциях тканей зуба.
Реализация функций:
1. Пульпа играет важную роль в физиологии твердых тканей зуба: через пульпу, наряду со слюной, осуществляется их питание.
2. Пульпа реализует гомеостатический контроль физико-химических параметров эмали зуба, и при ее удалении происходит снижение микротвердости эмали. Процессы де- и реминерализации твердых тканей зуба происходят почти параллельно, но в депульпированных зубах протекают быстрее, чем в зубах с сохраненной пульпой.
3. Кислотоустойчивость эмали зуба уменьшается после депульпирования. Функциональное состояние пульпы изменяет кислотоустойчивость эмали и вслед за снижением функциональной кислотоустойчивости возникает кариес.
4. В твердых тканях депульпированных зубов концентрация Са и Р уменьшается на 2-3%, а содержание О2 и его окисных форм увеличивается. Это в дальнейшем приводит к увеличению хрупкости зубов, лишенных пульпы.
ДЕНТИН
Дентин: продукт деятельности одонтобластов. Виды, состав и биохимические параметры дентина. Особенности органического матрикса интертубулярного дентина: отсутствие фибронектина, специфические гликопротеины (фосфофорин; дентинный сиалопротеин). Дентинная жидкость.
Дентин - основная масса зуба, представляет собой особого рода соединительную ткань. Минерализуется она по тому же типу, что и костная ткань.
Коронковая часть дентина покрыта эмалью, Корневая часть - цементом. Дентин построен из основного (или склеивающего вещества) и проходящих в нем трубочек.
По системе дентинных трубочек циркулирует дентиновая жидкость (дентиновая лимфа илизубной ликвор) и поступают питательные вещества (трофическая функция дентиновых канальцев). Количество дентиновой жидкости составляет около 0,01 мл на 1 г зуба. В ней содержатся калий, натрий, хлориды. Изменение состава дентиновой жидкости зависит от характера питания.
Дентиновые трубочки не подвергаются минерализации, поскольку отростки клеток покрыты снаружи слоем основного вещества из сильно гидратированных протеогликанов и не содержащего коллагена. Следовательно, здесь нет основы для образования центров кристаллизации. Этот слой основного вещества называют периплазматической зоной, её толщина невелика и составляет 150–200 нм. Сами канальцы имеют более широкий просвет: около тела клетки диаметр 2,0–3,0 мкм и у границы дентина с эмалью или цементом всего 0,5–1,0 мкм.
Дентин содержит: 72% неорганических веществ:
фосфата Са – Ca3(PO4)2,
карбоната Са – CaCO3,
фторида – CaF2,
28% органических веществ:
коллаген I типа (как в костях),
протеогликаны,
фосфопротеины и др.
В основном веществе содержится:
1) огромное количество минеральных солей, составляющих кристаллы гидроксиапатита. Гидроксиапатит дентина имеет повышенную плотность и твердость. Кроме гидроксиапатита есть немного аморфной (некристаллической) фазы
фосфатов и карбонатов Са. Дентин содержит в 3 раза больше Mg, чем кости.
2) коллагеновые фибриллы, собранные в пучки.
Следует отметить, что в предентине образуется избыток α1 цепей, что может быть причиной синтеза особого типа коллагена [α1(I)]3. Однако его не обнаруживают после прорезывания зубов.
Коллаген-I и дополнительный коллаген-V дентина синтезируется специализированными клетками - одонтобластами, которые располагаются в пульпе на границе с дентином. Они образуют тонкий слой, выстилающий полость зуба. Отростки клеток пронизывают дентин до сочленения его с эмалью, формируя дентиновые канальцы. В этих трубочках расположены не только отростки одонтобластов, но и нервные окончания из пульпы (структурная функция дентиновых канальцев).
Начинает синтезироваться мягкий предентин, который затем созревает и минерализуется. К моменту прорезывания получается зрелый дентин.
Первым еще на стадии преодонтобластов формируется плащевой дентин, который минерализуется по типу гомогенной нуклеации и далее при дозревании клеток и переходе к синтезу других видов дентина: интер-, перитубулярного постепенно отодвигается к границе с эмалью. В дентине, по сравнению с костной тканью, меньше остеопонтина и костного сиалопротеина. мало белков, содержащих γ-карбоксиглутаминовую кислоту (остеокальцин=костный гла-протеин и матриксный гла-протеин). Среди неколлагеновых белков дентина существенное место занимают специфические фосфогликопротеины. Особенности органического матрикса интертубулярного дентина: отсутствие фибронектина, специфические гликопротеины. которые образуются из одного белка-предшественника (фосфофорин – содержит RGD-центр для связи с клетками, богат АСП (40%) и СЕР (50%, почти все эстерифицированы фосфатом), придающими кислые свойства, для связывания с кальцием; дентинный сиалопротеин не содержит RGD-центра). Фосфопротеины появляются в предентине перед началом минерализации и располагаются на линии фронта минерализации.
Содержание протеогликанов в предентине выше, чем дентине (это связано с наличием в предентине ферментов, разрушающих протеогликаны).
В отличие от эмали клетки дентина функционируют на протяжении всей жизни зуба. Благодаря жизнедеятельности клеток формирование дентина может продолжаться после прорезывания зубов. При наличии жизнеспособной пульпы процесс образования дентина происходит в течение всего периода функционирования зуба. В результате дополнительно образуется вторичный дентин, который заполняет полость зуба и доставляет неудобства при лечебных вмешательствах. Вторичный дентин менее минерализован и содержит больше коллагена. Важное место в механизмах образования вторичного дентина отводят неколлагеновым белкам дентина (протеогликаны, фосфопротеины и др.). При кариесе в ходе формирования еще менее организованного третичного дентина их состав и количество также меняются в первую очередь.
Дентин проницаем для многих веществ, многие из них проникают в дентин главным образом через пульпу, в том числе ионы Са2+ при формировании дентина. После прорезывания зуба минерализация осуществляется, в основном через эмаль: Са-Р соединения поступают из слюнных мицелл.
При возникновении кариеса происходят большие биохимические изменения в органическом матриксе дентина. В участках, содержащих коричневый пигмент, резко увеличивается концентрация углеводов (до 4 %), в том числе гексозаминов. Часть коллагена дентина становится полностью резистентной к бактериальной коллагеназе – эта фракция особенно богата углеводами (до 14%). Понижается содержание пролина и оксипролина, а также аргинина, гистидина и оксилизина, уменьшается количество свободных эпсилон-аминогрупп.
Из твердых тканей зуба наиболее рано образуется дентин. Прилегающие к внутренним клеткам эмалевого органа (будущим амелобластам) соединительнотканные клетки зубного сосочка под индуктивным влиянием со стороны последних превращаются в дентинобласты, которые располагаются в один ряд наподобие эпителия. Они начинают формировать межклеточное вещество дентина - коллагеновые волокна и основное вещество, а также синтезируют фермент щелочную фосфатазу. Этот фермент расщепляет глицерофосфаты крови с образованием фосфорной кислоты и отщепляет эту кислоту от других органических фосфатов. В результате соединения фосфорной кислоты с ионами кальция формируются кристаллы гидроксиапатитов, которые выделяются между коллагеновыми фибриллами в виде матриксных пузырьков, окруженных мембраной. Кристаллы гидроксиапатита увеличиваются в размерах. Постепенно происходит минерализация дентина.