А. Строение оперона прокариот
Особенности организации генов про- и эукариот. Строение оперона
Практически любой ген несёт информацию о строении какой-либо РНК. Эта информация закодирована в определённой последовательности триплетов. Однако сам ген функционировать практически не может.
Необходим целый ряд дополнительных структур, зон или участков, которые не только включают и выключают работу гена, но и меняют интенсивность его работы в зависимости от нужд организма. К таким структурам относится множество разнообразных последовательностей ДНК, которые до настоящего времени не имеют общепризнанной классификации. Мы будем придерживаться наиболее простой (но далеко не полной) классификации. По ней дополнительные структуры подразделяются на два типа – регуляторные зоны и регуляторные гены. Регуляторные зоны это участки ДНК на которых не происходит синтез РНК, но которые служат местом связывания различных белков (или РНК). Эти последовательности часто называют регуляторными зонами, ( или регуляторные области, элементы, структуры, участки и др.). На регуляторных генах транскрибируется какая-либо РНК. Эта РНК может не кодировать белок, а осаждаться на регуляторной зоне гена. Но может и нести информацию о каком-либо белеке, тогда с регуляторной зоной связывается кодируемый РНК белок.
Таким образом, в настоящее время большинство учёных приходит к мнению, что наименьшей функциональной областью в ДНК является совокупность состоящая из структурного гена, регуляторных зон и регуляторных генов.
Сам ген представляет в основном кодирующую часть ДНК. Сразу же оговоримся, что у разных генов регуляторные области различны не только по строению, величине и другим параметрам, но и отличаются по пространственному положению относительно гена (или генов), функцию которого они курируют. Понятно, что рассмотреть строение всех известных генов и их регуляторных областей со всеми индивидуальными особенностями в нашем пособии невозможно. Поэтому приведём структуру некого гипотетического гена и его регуляторных областей, обозначив у них те участки, которые наиболее часто встречаются.
Кроме того, мы рассмотрим строение не всех структурных генов, а тех, с которых транскрибируется иРНК, несущая информацию о структуре белка. Это важно помнить, т.к. строение других генов (с которых транскрибируются тРНК, рРНК или регуляторных генов несколько иное).
Как мы отмечали раньше, для нормального функционирования гена необходимы регуляторные зоны. Регуляторных генов в этом разделе касаться не будем.
У прокариот регуляторные зоны «обслуживают» несколько генов. Эти гены вместе с регуляторными элементами носят название оперон. Таким образом, оперон состоит из двух функционально различных участков: (см. рис. 18, А).
- Кодирующего участка, который содержит несколько структурных генов.
- Регуляторной зоны , которая включает следующие участки:
а. Стартовый кодон – сайт (место) инициации транскрипции.
б. Терминатор – сайт конца транскрипции.
в. Лидирующую область.
г. Трейлерную область.
д. Промотор.
е. Оператор.
ж. Активатор
з. Спейсеры.
Все эти участки представлены на рисунке 18 (Б,В,Г).
Как отмечали выше третий элемент – регуляторные гены, которые непременно входят в состав наименьшего функционирующего участка, в этом разделе рассматриваться не будут.
Оперон

Регуляторная зона Кодирующая область (Гены)

 А 5’ 3’
А 5’ 3’
 |
Регуляторная зона Ст. 1 Ст. 2 Ст. 3

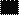


 Б5’ 3’
Б5’ 3’



 Сайт начала транскрипции Терминатор
Сайт начала транскрипции Терминатор








 В5’ 3’
В5’ 3’




Лидерная последовательность Трейлерная последовательность.




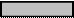

 Ак. Пр. Оп. К о д и р у ю щ а я о б л а с т ь
Ак. Пр. Оп. К о д и р у ю щ а я о б л а с т ь
Г5’ 3’
Рис. 18. Схема регуляторных и кодирующих элементов оперона прокариот. На рисунке от А до Г повышается детализация строения оперона, его регуляторной и кодирующей области. Ст.1,2,3 – структурные гены. Ак. – активатор, Пр. – промотор, Оп. – оператор.
Кодирующая область (собственно ген) начинается с сайта инициации (стартовый кодон). С этого участка РНК-полимераза, проходя через структурный ген, начинает синтезировать РНК. Заканчивается кодирующая область участком, который называется терминатор(рис. 18, Б). Подходя к нему, РНК-полимераза заканчивает транскрипцию и сходит с нити ДНК. Терминирующая область у многих генов имеет различное строение. Отметим два из них.
1. Чаще всего в терминирующей области располагается один из бессмысленных кодонов (УАА, УАГ, УГА), не кодирующий ни одну аминокислоту. Обнаружив эту последовательность РНК-полимераза прекращает синтез РНК.
2. Сигналом к окончанию транскрипции могут быть определённые короткие последовательности ДНК (не бессмысленные кодоны), которые располагаются в зоне окончания синтеза РНК. К этим последовательностям прикрепляется белок, который и прекращает транскрипцию.
В последнее время обнаружили, что в зоне терминатора ДНК может формировать шпильки, которые и приводят к окончанию транскрипции.
Область, располагающаяся между сайтом инициации и терминации, транскрибируется как одна нить РНК и носит название единица транскрипции. У прокариот единица транскрипции, как правило, содержит последовательности, кодирующие не один, а несколько типов белков или РНК, т.е. содержит несколько структурных генов (рис. 18, Б). Все они имеют одну регуляторную область и контролируют синтез ферментов одного биохимического цикла (на рисунке они обозначены как Ст1, Ст2 Ст3).
Кроме перечисленных регуляторных зон обнаружено, что перед стартовым кодоном и терминаторомрасполагаются небольшие участки ДНК, которые соответственно носят название лидерные и трейлерные области (или последовательности). Лидерная область включает или отключает транскрипцию иРНК, трейлерная принимает участие в «созревании» иРНК (рис. 18,В).
Особенностью лидерного участка является то, что он транскрибируется, т.е представлен в молекуле иРНК. Но этот участок в рибосомах не транслируется, т.е. он не представлен аминокислотной последовательностью в белке. Более тщательные исследования показали, что лидерная последовательность обладает уникальной способностью приобретать форму шпильки в том случае, когда транскрипция данного гена клетке не нужна. Например, при отсутствии субстрата, нет необходимости транскрибировать иРНК и транслировать с неё фермент, расщепляющий субстрат. Поэтому довольно часто отсутствие субстрата провоцирует образование в лидерной последовательности шпильки и синтез иРНК не начинается.
Трейлерная последовательностьтранскрибируется на иРНК и является сигналом для формированию поли(А) хвостика при «созревании» про-иРНК (см. далее).
Начиная с 5’ конца, по направлению к 3’ концу располагаются – активатор, промотор и оператор(рис. 18, Г). К промотору присоединяется РНК-полимераза. Активатор и оператор регулируют активность гена. Так к активатору присоединяется белок, способный облегчить присоединение фермента к промотору или, наоборот, затормозить этот процесс. На операторе также осаждается белок, который может блокировать работу РНК-полимеразы.
Ещё раз подчеркнём важную особенность функционирования оперона у прокариот:- одна регуляторная область оперона (куда входят активатор, промотор, оператор, стартовый кодон и др.), как правило, обслуживает несколько структурных гена. Причём, между последними располагаются последовательности ДНК не несущие никакой информации. Эти последовательности называютспейсерами (см. рис. 19).
РОСт.1 Ст.2 Ст.3 Ст.4

С п е й с е р ы Терминатор
Рис. 19 Кодирующая область оперона, включает 4 структурных гена (Ст.1,Ст2, Ст3, Ст4), которые разделены спейсерами. Обслуживаются они одной регуляторной областью (на рисунке она обозначена как РО).
Несколько слов о правилах обозначения генов. В разделе 1 мы уже объяснили, на каком принципе основано обозначение концов ДНК, если последняя представлена на графике в виде линейного изображения.
На рисунке 18 изображена одна нить ДНК, которая маркирована цифрами 5’( начало отрезка) и 3’(конец отрезка). Эти обозначения общеприняты и облегчают понимание генетических процессов происходящих на гене (транскрипции и трансляции), которые имеют выраженную направленность. Однако в обозначении направленности гена имеются некоторые особенности, без знания которых определить начало и конец отрезка ДНК сложно. Из школьного курса известно, что транскрипция РНК происходит на двухнитчатой ДНК только с одной её нити.
Эта нить ДНК называется матричной (этот термин употребляется чаще всего), антисмысловой, незначащей, не кодирующей, кодогенной и т.д. Понятно, что синтезированная с этой нити РНК будет комплементарна ей. Однако полное совпадение последовательностей нуклеотидов у вновь синтезированной РНК будет не с нуклеотидами матричной цепи, а с противоположной (второй, оппозитной) нитью ДНК, которая называется смысловая, не матричная, значащая, кодирующая, не кодогенная и т.д.
Транскрипция с матричной цепочки ДНК идёт в направлении от 3’ конца к 5’. Понятно, что противоположная цепочка ДНК будет иметь направление 5’ – 3’. Это направление и принято обозначать на рисунках. Поэтому необходимо помнить, что на рисунках принято обозначать цифрами не ту цепочку ДНК, с которой синтезируется (транскрибируется) РНК, а противоположную – смысловую (см. рис. 20).


 5’ А Т А Т Г Ц А Т Г Ц 3’ Смысловая нить
5’ А Т А Т Г Ц А Т Г Ц 3’ Смысловая нить





 А ДНК
А ДНК


 3’ Т А Т А Ц Г Т А Ц Г 5’ Антисмысло-
3’ Т А Т А Ц Г Т А Ц Г 5’ Антисмысло-
вая нить ДНК, с
которой синтези-
руется иРНК
 Б РНК 5’ А Т А Т Г Ц А Т Г Ц 3’ РНК синтезирован-
Б РНК 5’ А Т А Т Г Ц А Т Г Ц 3’ РНК синтезирован-
ная с антисмысло-
вой нити ДНК
 В ДНК 5’ А Т А Т Г Ц А Т Г Ц 3’Так на рисунках обо-
В ДНК 5’ А Т А Т Г Ц А Т Г Ц 3’Так на рисунках обо-
значается отрезок ,
ДНК. Последователь-
ность нуклелтидов-
в ней аналогична по-
следоватнельности
их в иРНК и антисмы
словой цепи ДНК.
Рис. 20. Схема транскрипции и правила обозначения направленности ДНК. А – отрезок молекулы ДНК, Б – синтезированная РНК на атисмысловой нити, В – цепь ДНК на рисунке обозначена так же, как и смысловая цепь ДНК