СТРУКТУРА БИОЦЕНОЗА
Чем больше отклоняются от нормы (оптимума) условия существования в пределах биотопа, тем беднее видами становится биоценоз, но при этом тем большей численностью особей будет представлен каждый вид. Следовательно, число особей и число видов находятся в обратной зависимости.
Чем разнообразнее условия существования в пределах биотопа, тем больше количество видов может существовать в данном биоценозе.
Эти два пункта дополняют и расширяют биоценотические принципы П.Жаккара(1928), согласно которым:
1. Видовое богатство территории пропорционально разнообразию экологических условий.
2. Биоценотическое разнообразие, зависящее от условий биотопа, климата, почвы, рельефа и т. д., нарастает с увеличением площади (пространства) и снижается по мере увеличения однообразия условий (исключение: зависимости от экстремальных показателей температуры, аридности или концентрации солей).
Для того, что бы лучше уяснить эти принципы следует обратиться к строению, т.е. к структуре биоценозов.
Получив представление о составе биоценозов, что в большинстве случаев требует натурных исследований, можно приступать к изучению структуры. Структура биоценоза – это взаимное расположение и соподчинение составляющих его компонентов, входящих в него систем, и все это – в динамике, с изменениями, как в пространстве, так и во времени. Нередко к структуре относят и различные взаимодействия между этими компонентами и системами, однако мы эти взаимодействия будем рассматривать в пределах функционирования биоценозов.
Пространственная структура экосистем
В любой, даже самой однородной системе можно при желании выделить хотя бы слега различные детали. Вопросов с подобным разграничением для биоценозов не возникает – любой из них, от самого мелкого, до самого крупного представляет собой некую совокупность неоднородностей, что определяется целым рядом причин. Так в лесу кроме древостоев, присутствуют поляны, редины, прогалины, болота, просеки, опушки и т.д. При этом сами древостои различаются по породному составу, сомкнутости, возрасту и др. Далее, однопородные древостои оказываются неодинаковыми по ярусам, составу напочвенной растительности, заболоченности и т.д., и т.д. Все эти неоднородности являются структурными элементами биоценозов.
Хотя наличие или отсутствие в экосистеме того или иного структурного элемента, а также его пространственное расположение в каждом конкретном случае может быть случайным, однако набор этих случайностей в рамках той или иной экосистемы имеет, как правило, закономерный характер. Поэтому при изучении пространственной структуры биоценоза, кроме вполне очевидной задачи описания составляющих его разностей, исследователь должен ответить на вопрос о причинах, повлекших наблюдаемое размещение элементов структуры. Обычно эти элементы представлены биоценозами более низких уровней.
Вопрос об уровнях биоценотической организации, или о масштабах исследования экосистем возникает и будет возникать постоянно. Выделять тот или ной уровень организации целесообразно в том случае, если на нем возникают новые свойства, отсутствующие у систем нижележащего уровня. Этот так называемый принцип эмерджентности. Эмерджентность (emergence – возникновение, появление) – одно из важнейших понятий теории систем. Синоним этого термина – системный эффект. Он означает наличие или возникновение у какой-либо системы особых свойств, не присущих её подсистемам, блокам, равно как и сумме элементов, не связанных особыми системообразующими связями. Это – несводимость свойств системы к сумме свойств её компонентов. Например, феномен жизни возникает на клеточном уровне (молекулы, из которых состоят клетки – НЕ живые организмы). Другой феномен – потенциальное бессмертие или, как минимум огромное долгожительство – возникает на популяционном уровне (организмы, из которых состоят популяции или виды, им не обладают). В биологии и экологии понятие эмерджентности можно выразить так: одно дерево — не лес, скопление отдельных клеток — не организм. Например, свойства биологического вида или биологической популяции не представляют собой свойства отдельных особей. Поэтому такие понятия как «рождаемость» и «смертность» НЕприменимы к отдельной особи, но вполне корректны по отношению к популяции или виду в целом. В теории эволюции эмерджентность выражается как возникновение новых функциональных единиц системы, которые не сводятся к простым перестановкам уже имевшихся элементов. Например, эмерджентным свойством почвы является плодородие, это свойство отсутствует у грунтов, на которых развивается почва и из которых она частично состоит.
Итак, составив таблицы с данными о видовом составе и обилии каждого вида живых организмов в биоценозе можно попытаться провести определенные манипуляции. То есть группировки. Например, объединить некоторые виды в группы по их таксономическому составу и обилию. Наиболее примитивное объединение видов по обилию – это выяснение степени доминирования – отношения(обычно в процентах)количества экземпляров данного вида к суммарному количеству особей всех видов в биоценозе. С помощью этой информации для каждого яруса или синузии удается провести сортировку на виды доминанты и второстепенные виды. Во многих биоценозах удается выделить один или несколько видов-доминантов – это виды, определяющие его облик и занимающие господствующее положение в определенном ярусе биоценоза. Обычно наземные биоценозы называют по доминирующим видам: ельник, дубрава или дубняк, сфагновое болото, ковыльно-типчаковая степь и т.д. Второстепенные виды – малочисленные и редкие также очень важны в сообществе. Их присутствие – это необходимые условия устойчивого развития экосистемы.
Общий принцип варьирования количества доминирующих видов по разным экосистемам формулирует правило Р.Крогеруса (1932): в биотопах с экстремальными условиями доминируют строго специализированные виды (для растений – стенотопные) с относительно большим количеством особей. Обратной стороной этого правила является другой широко известный факт: в обычных и особенно в благоприятных для жизни условиях может сосуществовать большое количество видов, с небольшой численностью каждого. При этом выделить доминантов в таких сообществах бывает совсем не просто.
В том случае, когда кроме видового состава и обилия мы располагаем некими данными об изменениях параметров среды (в пределах данного биоценоза), можно провести градиентный анализ сообщества. Его суть сводится к размещению на двухмерном графике точек, в которых обилие конкретного вида характеризуется разными значениями. Этими значениями определяется положение данной точки относительно ординаты. Осью Х характеризуется какой-то один градиент среды: влажность почвы, или ее механический состав, глубина снежного покрова и т.д. Соответствие данного фактора среды его реальным условиям в конкретной точке определяет положение этой точки относительно горизонтальной оси. Линия, которой соединяются все точки одного вида организмов, показывает не только реакцию конкретного вида на данный фактор среды, но и дает некое представление о пространственном расположении этого вида. Обычно на один график (один фактор среды) наносятся показатели обилия разных видов, что дает возможность оценить их взаимное расположение (рис. 2).
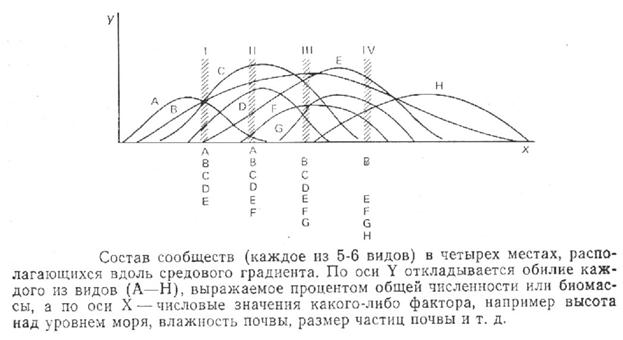 Рисунок 2
Рисунок 2
По взаимному расположению видов их обычно удается объединить в группы, которые, вероятно, будут занимать разные биотопы. Однако проведение четких границ в большинстве случаев будет затруднено, что укажет на сложность выявления пространственной структуры биоценоза.
Чем круче градиент (сильнее изменение значений) данного фактора среды, чем более четкими окажутся границы размещения разных видов. В экстремальных условиях (например, в аридных или в холодных) градиенты бывают особенно крутыми. В таких случаях доминирующие виды часто своим распределением четко обозначают границы сообществ, что выражается в относительной легкости, с которой удается распознавать и классифицировать сообщества экстремальных условий. В умеренных зонах сделать это гораздо сложнее.
Главный недостаток градиентного анализа – его субъективность. Во-первых, потому, что исследователь подбирает для анализа только те градиенты среды, которые, по его мнению, важны для организмов. Кроме того, корреляционная связь не всегда отражает жизненную необходимость организмов в данном факторе. Градиентный анализ – лишь ступень в изучении структуры сообщества.
Метод ординации позволяет группировать данные независимо от мнения исследователя о сочетаемости видов или о переменных среды, определяющих распределение видов в пространстве. Его суть – размещение в неком пространстве эмпирических точек с данными о видовом составе и обилии видов (для чего используются методы класстерного и дискриминантного анализов).
Оси этого пространства выводятся математическим путем только на основании состава и обилия видов в эмпирических точках. Но одновременно эти же оси соответствуют градиентам каких-то факторов, которые действительно влияют на организацию сообществ. Поэтому следующий шаг – это идентификация этих осей, т.е. распознавание в них экологически знàчимых переменных среды (эта процедура проводится с помощью анализа главных компонент). Заранее установить какие факторы окажутся знàчимыми невозможно. Невозможно и задним числом установить знàчимый фактор, если он пропущен при сборе данных.
Результаты ординации более объективны. К тому же они подчеркивают, что при заданном сочетании условий среды, возникает вполне определенная ассоциация видов. Следовательно, при изучении сообществ эколог имеет дело не с произвольным набором видов, а с чем-то большим.
Классификация, в противоположность ординации, исходит из допущения, что сообщества относительно дискретны. Такой взгляд позволяет выделять группы родственных сообществ, аналогично, например, таксономической классификации: в систематике близкие виды объединяются в роды, близкие роды в семейства и т.д. Аналогичным образом сообщества со сходным видовым составом и разнообразием объединяются в группы, которые в случае необходимости, могут объединяться и дальше по признакам сходства. Основой для таких группировок обычно являются результаты кластерного анализа.
И ординация, и классификация позволяют обнаружить упорядоченность, или структуру, в совокупности сообществ, не выбирая предварительно какого-либо важнейшего (ведущего) фактора среды, как в случае градиентного анализа.
Результаты многих исследований, выполненных с помощью представленных здесь методов изучения пространственной структуры сообществ, показывают, что некий вид из той или иной ассоциации при лишь немного отличных условиях среды вполне может оказаться и в другом месте, среди других видов. Такое положение дел обусловлено индивидуальными свойствами видов и вмещающей их среды:
1. Все организмы устойчивы в лишь неком диапазоне условий среды.
2. Разные виды имеют разные диапазоны устойчивости.
3. В пределах вида особи различаются между собой по экологическим требованиям.
4. Сами условия среды меняются в пространстве по градиентам, т.е. постепенно.
Таким образом, четкие границы между сообществами НЕвозможны за исключением случаев резкой, ступенчатой смены условий среды. Этот вывод позволяет понять, почему границы, используемые в биогеографии для выделения природных зон или биомов в значительно степени произвольны, да и окончательного согласия о количестве этих крупных экосистем пока не достигнуто.
Именно индивидуальные особенности видов и конкретных условий среды приводят к существенным внешним различиям биоценозов, среди которых следует упомянуть два крайних варианта: простые и сложные биоценозы.
Сложными биоценозами называются многоярусные сообщества, представленные большим количеством видов растений, животных и микроорганизмов, функционально связанных между собой многообразными взаимоотношениями. Именно такая сложность определяет их наибольшую устойчивость к неблагоприятным внешним (экзогенным) и внутренним (эндогенным) воздействиям. Исчезновение какого-либо вида существенно не отражается на судьбе таких биоценозов, т.к. его индивидуальная роль в жизни всего сообщества, как правило, весьма локальна и в большинстве случаев в той или иной мере дублируется другими видами (возможно несколькими). В таких биоценозах происходят лишь незначительные структурные перестройки, при которых популяции одного и даже нескольких видов могут замещаться экологически близкими видами, а стабильность сообщества определяется количественной регуляцией численности одних видов другими. Так, в самых сложных биоценозах тропических лесов никогда не наблюдаются вспышки массового размножения отдельных видов.
Простые биоценозы всеми этими особенностями НЕ обладают. Они характеризуются упрощенной ярусностью и видовым составом (например: кустарничковые тундры – один растительный ярус с 2-3я синузиями, лишайниковые сосняки северной тайги – 2 яруса с 2-3я синузиями). Для большинства простых биоценозов обычны «вспышки» численности отдельных видов-доминантов. Так, в тундрах обычны резкие колебания численности леммингов с огромной амплитудой пиков и депрессий. Типичны также падения и взлеты численности песцов, питающихся леммингами; сильно колеблется и численность северных оленей, оказывающих существенное влияние на растительный покров. Главные причины таких колебаний – отсутствие (или недостаток) видов-«дублеров», которые при необходимости могли бы заменить основные пищевые объекты и выступить в качестве «буферного» корма для фитофагов и/или хищников.