Основные механизмы регуляции функционирования оперона прокариот
Гены-регуляторы являются важными элементами оперона. Под действием клеточных факторов они обусловливают синтез регуляторных белков. Такие белки, соединяясь с определенными нуклеотидными последовательностями ДНК (оператором), могут способствовать или препятствовать присоединению РНК-полимеразы к промотору. В случае если белок регулятор не дает возможность ферменту присоединяться к промотору (слайд 2Б), он называется репрессором. В этом случае осуществляется негативный контроль транскрипции со стороны гена-регулятора. В случае если белок-регулятор способствует присоединению РНК-полимеразы к промотору и началу процесса транскрипции, его называют белком-активатором, и осуществляется позитивный контроль со стороны гена-регулятора (слайд 2В).
В процессах регуляции экспрессии генов принимают участие также вещества небелковой природы (эффекторы), взаимодействующие с белками-регуляторами и изменяющими их способность связываться с опероном. Например, конечный продукт метаболической цепи. В зависимости от результатов такого воздействия среди эффекторов различают индукторы, способствующие транскрипции, и корепрессоры, препятствующие ей.
Функционирование лактозного оперона Е.соli
 В своих экспериментах Джакоб и Моно обнаружили, что добавление лактозы к культуре Е.соli индуцирует образование сразу трех белков-ферментов: галактозидазы, пермеазы и трансацетилазы, необходимых клетке для расщепления лактозы до глюкозы и галактозы. Гены, кодирующие эти ферменты, соседствуют друг с другом в хромосоме. Их назвали структурными генами, или цистронами (слайд 3).
В своих экспериментах Джакоб и Моно обнаружили, что добавление лактозы к культуре Е.соli индуцирует образование сразу трех белков-ферментов: галактозидазы, пермеазы и трансацетилазы, необходимых клетке для расщепления лактозы до глюкозы и галактозы. Гены, кодирующие эти ферменты, соседствуют друг с другом в хромосоме. Их назвали структурными генами, или цистронами (слайд 3).
Схема функционирования лактозного оперона E.coli
Они одновременно транскрибируются РНК-полимеразой в длинную одиночную мРНК, которая имеет кодоны для всех трех ферментов. мРНК, транскрибируемая из нескольких генов, называется полицистронной. Способность к транскрипции этих цистронов контролируется участком молекулы ДНК — оператором. Операторный локус — это определенный участок оперона, имеющий определенную последовательность нуклеотидов, длиной 27 пар оснований. Этот сегмент ДНК располагается в области промотора, к которому перед началом транскрипции присоединяется РНК-полимераза. Промотор расположен перед началом первого структурного гена β-галактозидазы. Цистрон «синтезирует» мРНК, когда оператор включен и прекращает синтез, когда он «выключен». Оператор включается или выключается белком, называемым репрессором. Его синтез контролируется регуляторным геном. Репрессор либо связывается с оператором, подавляя его активность, либо не связывается с ним, позволяя проявление активности структурных генов. Таким образом, репрессор является негативным регулятором.
На слайде 3 представлена схема функционирования лактозного оперона. Репрессия (I). При отсутствии индуктора (лактозы), репрессор связывается с оператором и блокирует транскрипцию, в этом случае ферменты не образуются. Индукция (II). Лактоза индуцирует транскрипцию генов, т.к. инактивирует репрессор. К промотору присоединяется РНК-полимераза и начинается транскрипция генов А, В и С. В результате образуются необходимые в данных условиях ферменты. Синтез Ферментов может не только индуцироваться, но и подавляться.
Например, в результате какой-то цепи реакций в клетке образуется конечный продукт Д в большем, чем это необходимо клетке, количестве. Это может нарушить нормальный ход реакций обмена, поэтому в клетке возникает необходимость остановить данный процесс. Тогда вещество Д вступает в реакцию с соответствующим белком-репрессором и переводит его в активное состояние. После этого происходит присоединение репрессора к оператору, тем самым выключается вся система оперона и синтез ферментов прекращается. В данном случае, торможение синтеза производится конечным продуктом, образующимся в результате реакции. Такой механизм действия называется регуляцией по принципу обратной связи (слайд 3-I).
Описанный механизм регуляции имеет большое приспособительное значение. Например, согласно имеющейся генетической программе, кишечная палочка может синтезировать несколько десятков ферментов, расщепляющих различные вещества, так как состав среды, окружающей бактерии, очень изменчив. В этих условиях постоянное образование всего набора ферментов было бы неэкономичным для клетки, так как нецелесообразно продуцировать одновременно 60-80 ферментов, из которых в данных условиях среды могут понадобиться лишь 6-8. Поэтому синтез ненужных в этих условиях ферментов блокирован, а клетки образуют лишь несколько необходимых белков. В природе отбор идет по принципу наибольшей экономии материи и энергии, поэтому клетки, функционирующие более экономично, лучше приспосабливаются и быстрее размножаются. Это и привело к совершенствованию системы регуляции метаболизма.
Особенности экспрессии генов у эукариот
Принципы регуляции экспрессии одинаковы как у прокариот, так и у эукариот. Однако эукариоты, особенно многоклеточные, — более сложные организмы и экспрессия их генов сложнее и несколько отличается деталями. В частности, можно отметить следующие особенности экспрессии у эукариот:
1. Геном высших эукариот значительно сложнее. Например, гаплоидный геном человека имеет приблизительно 35 тысяч генов, расположенных в 24 (22 аутосомы + X и Y) хромосомах. А у прокариот имеется
только одна хромосома и несколько сотен генов.
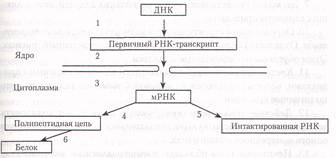
2. В клетках эукариот ядерная оболочка пространственно разделяет
процессы транскрипции и трансляции, хромосомы находятся в ядре,
а рибосомы — в цитоплазме. Экспрессия генов у эукариот включает
гораздо большее число этапов. Поэтому у них есть ряд механизмов регуляции, отсутствующих в клетках прокариот, например, процессинг.
3. На экспрессию эукариотических генов оказывает влияние амплификация генов. Это многократное увеличение числа копий одинаковых
генов с целью интенсификации синтеза молекул, нужных в определенный момент времени. Например, повторяющиеся последовательности
ДНК включают сотни копий генов рРНК и тРНК.
4. В отличие от прокариот, структурные гены эукариот имеют продолжительные последовательности азотистых оснований, которые не
кодируют аминокислот — интроны. Между ними расположены последовательности, которые кодируют аминокислоты — экзоны. Транскрибируемая из гена РНК имеет как интроны, так и экзоны. Она называется
про-мРНК. Ее интронные области удаляются нуклеазами, а несущие
информацию участки — экзоны соединяются вместе. Процесс обработки
про-мРНК и превращение ее в мРНК известен как процессинг.
5. У эукариот не установлено полной оперонной организации генетического материала. Гены ферментов определенной метаболической
цепи могут быть расположены в различных хромосомах. Они обычно
не имеют общей системы регуляции в виде гена регулятора, оператора и промотора. Поэтому синтезируемые в ядрах эукариот мРНК моноцистронны. Регуляция активности генов у эукариот сложнее, т.к. в этом процессе участвуют сразу несколько генов-регуляторов, то есть регуляция транскрипции эукариот является комбинативной. Например, у эукариот на молекуле ДНК имеется специальная область около промотора. Эта область имеет около 100 пар специальных нуклеотидов (препромоторный элемент). К этому участку молекулы присоединяется особый белок — фактор транскрипции. Это обеспечивает успешное присоединение РНК-полимеразы II к промотору.
6. Еще одна особенность генома эукариот — наличие специальных
«усиливающих» сегментов ДНК — энхансеров. Они могут располагаться на большом расстоянии от промотора. Их функцией является
участие в регуляции активности структурных генов. В свою очередь
и препромоторный элемент, и энхансер регулируется соответствующими регуляторными белками. Некоторые белки-регуляторы обладают
координирующим влиянием на активность многих генов, то есть они
обладают плейотропным действием.
7. Геном эукариот подчиняется регуляторным воздействиям со
стороны эндокринной системы организма. Многие гормоны являются
индукторами транскрипции. В первую очередь это относится к стероидным гормонам, которые обратимо связываются с белками-рецепторами,
переносящими их в ядро. Такой комплекс связывается со специфическим участком хроматина, ответственным за регуляцию генов. Например, действие тестостерона активизирует гены, определяющие развитие
организма по мужскому типу.
8. Еще одна особенность регуляции активности генов эукариот
связана с образованием хроматина — комплекса ДНК с белками.
В таком виде гены в составе нуклеосом неспособны к транскрипции.
Поэтому необходимым условием является частичная декомпактизация хроматина и ослабление связей с гистоновыми белками. Однако
полная нуклеосомная организация хроматина в ходе транскрипции не
утрачивается.
9. Контроль экспрессии генов у эукариот осуществляется также на
стадии трансляции, например, путем воздействия на фактор инициации
трансляции. Поэтому даже при наличии в цитоплазме мРНК синтез на
ней может не происходить.
10. Некоторые гены эукариот многократно повторены, а определенные же участки ДНК вообще не играют генетической роли, например,
сателитная ДНК. Геном эукариот «избыточен». Например, у человека
функционирует 2-3 % из 35 000 всех генов. Кроме этого, в геномах эукариот содержатся последовательности, повторяющиеся несколько десятков, сотен и даже миллионов раз. Среди повторяющихся последовательностей имеются элементы с непостоянной локализацией. Их называют транспозонами, или мобильными элементами. Повторяющиеся последовательности выполняют разнообразную биологическую роль: регуляция воспроизведения ДНК, участие в кроссинговере, обозначение границы между экзонами и нитронами и др.
11. Регуляция экспрессии может осуществляться на стадии пострансляционных изменений (модификация). Например, для образования
активной формы белкового гормона инсулина из молекулы проинсулина
вырезаются две цепочки, которые затем сшиваются дисульфидными
связями.
12. В процессе синтеза одного белка участвует не один, а много
генов. В частности, один или несколько структурных генов, образующих мРНК, примерно 20 генов тРНК, необходимых для переноса
20 различных аминокислот (мономеров белков) и несколько генов
рРНК, образующих рибосомы. Функционируют также много других
структурных генов: различных ферментов процессинга, трансляции,
модификации и т. д.
Итак, очевидно, что экспрессия гена в признак эукариот — очень сложный процесс, который регулируется на многих уровнях (слайд 4), в том числе другими генами. Таким образом, любой признак эукариотического организма является полигенным.
 Схема, иллюстрирующая шестъ уровней, на которых может осуществляется контроль экспрессии генов у эукариот:
Схема, иллюстрирующая шестъ уровней, на которых может осуществляется контроль экспрессии генов у эукариот:
1 — контроль на уровне транскрипции;
2 — контроль на уровне процессинга;
3 — контроль на уровне транспорта;
4 — контроль на уровне трансляции;
5 — контроль на уровне деградации;
6 — контроль на уровне модификации
Современное состояние теории гена
В результате исследований элементарных единиц наследственности сложились представления, носящие общее название теории гена. Основные положения этой теории следующие:
1. Ген - участок молекулы ДНК, имеющей определенную последовательность нуклеотидов. Представляет собой сложную функциональную единицу наследственной информации, состоящую из различных
функциональных сегментов.
2. Разные гены имеют разный качественный и количественный со
став нуклеотидов.
3. Каждый ген имеет определенное место (локус) в хромосоме.
4. Гены способны к рекомбинации (в процессе кроссинговера) и му
тации, что обеспечивает изменчивость.
5. В хромосоме есть гены мРНК (структурные гены), гены рРНК
и гены тРНК.
6. Среди структурных генов есть регуляторные гены, продукты
которых регулируют работу других структурных генов.
7. Ген не принимает непосредственного участия в синтезе белков, он
является матрицей для образования посредников — различных молекул
РНК, непосредственно участвующих в синтезе.
8. Количество генов может удваиваться в процессе репликации,
а затем распределяться в дочерние клетки в результате митоза или
мейоза.
9. Ген может существовать в виде нескольких аллелей, определяющих варианты признаков.
10. Определенный структурный ген кодирует синтез одного полипептида. Отдельный белок может обусловливать определенный признак.
Этим обусловлены моногенные признаки.
11. Клетка, орган или организм обладают многими сложными при
знаками, которые слагаются из взаимодействия многих генов - это
полигенные признаки.
12. Действие гена строго специфично, т. к. ген может кодировать
только одну аминокислотную последовательность и регулирует синтез
одного конкретного полипептида.
13. Некоторые гены обладают плейотропностъю действия (см.
главу 5), определяя развитие сразу нескольких признаков.
14. Дозированностъ действия гена заключается в зависимости интенсивности проявления признака (экспрессивность) от количества определенного аллеля. Например, многие заболевания в гетерозиготном состоянии проявляются слабее, чем в гомозиготном.
15. На активность гена может оказать влияние как внешняя, так
и внутренняя среда.
16. Конститутивные гены - это гены, которые постоянно экспрессируются, так как белки, которые они кодируют, необходимы для
постоянной клеточной деятельности, обеспечивают синтез белков
«домашнего хозяйства» - белков рибосом, цитохромов, ферментов
гликолиза, переносчиков ионов и др. Эти гены не требуют специальной
регуляции.
17. Неконститутивные гены - это гены обычно неактивные, но
экспрессирующиеся только тогда, когда белок, который они кодируют,
нужен клетке. Эти гены регулируются клеткой или организмом. Эти
белки обеспечивают дифференцировку, специфичность структуры
и функций каждой клетки.
18. Молекулы ДНК способны к репарации, поэтому не всякие повреждения гена ведут к мутациям.
19. Генотип, будучи дискретным (состоящим из отдельных генов),
функционирует как единое целое.
Взаимосвязь между генами, ферментами и признаками
Тесная взаимосвязь между генами и ферментами была известна задолго до того, как был установлен механизм экспрессии гена. Она была обнаружена в 1909 г. британским врачом А.Гарродом. Он предположил, что заболевания, наследуемые потомками от родителей, могут быть вызваны недостатком ферментов, которые катализируют превращение одного метаболического вещества в другое. Он назвал такие заболевания «врожденными ошибками метаболизма». Недостаток фермента связан с отсутствием нормальной аллели гена, который контролирует синтез данного фермента. Эта концепция в 1945 г. была положена в основу гипотезы Дж. Бидла и Э. Татума: один ген — один фермент. Это положение утверждает, что каждый ген контролирует синтез своего белка. Но некоторые белки содержат более чем одну полипептидную цепь, например, гемоглобин состоит из 2-х α- и 2-х β-полипептидных цепей. Значит, образование сложного белка требует участие нескольких генов. Таким образом, гипотеза была уточнена: один ген — одна полипептидная цепь. Результатом участия белков в метаболизме является развитие признака или признаков организмов. Весь процесс от биосинтеза РНК и до проявления свойств белков можно представить в виде следующей цепи событий:


 ДНК про-мРНК мРНК полипептидная цепь белок
ДНК про-мРНК мРНК полипептидная цепь белок
 признак.
признак.
В результате «работы» генов образуются функционально активные белки или их гликолипидопротеиновые комплексы, которые включаются в метаболизм, либо секретируются (выводятся) из клеток. С током крови они поступают к другим органам и выполняют там определенную роль: ферментативную, регуляторную или структурную. Например, гормон инсулин синтезируется как более крупная молекула — проинсулин, не имеющий активности. В процессе модификации в аппарате Гольджи ненужная часть молекулы отцепляется, а две образующиеся полипептидные цепи сшиваются ферментами. Гормон приобретает определенную структуру и функциональную активность. Его активность обеспечивает наличие признака организма — постоянство глюкозы в крови человека.