Молекулярно-биохимическая организация саркомера. Основные, регуляторные и вспомогательные белки мышечной ткани.
Рис. 2.3.6. Расположение митохондрий в мышце млекопитающих
Рис.2.3.3. Взаиморасположение актиновых и миозиновых фибрилл
Рис. 2.3.1. Микрофотография (скан.эл.микр.) и фото саркомера скелетной мышцы.
Рис. 2.2.3 Строение мышцы. В центре виден центральный пучок из нервов, кровеносных сосудов и интафузальных волокон (мышечное веретено - 10) (по R.Krstic, 2001)
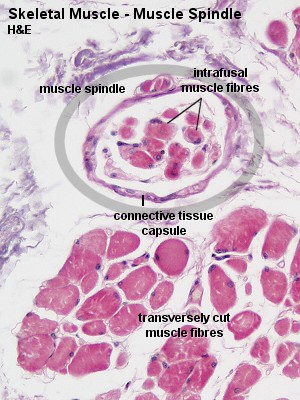
Рис. 2.2.4. Гистология участка мышцы позвоночных, включая экстрафузальные и интрафузальные мышечные волокна. Участок «мышечного веретена», содержащий интрафузальные мышечные волокна обведен кругом.

Рис. 2.2.5. Схема строения «мышечного веретена» и система его иннервации (по: Гистология, 2001)
2.3. Саркомер – как основная морфо-функциональная единица мышечного волокна.
Саркомер является структурной единицей миофибрилл поперечно-полосатого мышечного волокна соматической (скелетной) мышцы позвоночных , а также ряда мышц беспозвоночных животных и миофибрилл кардиомиоцитов. (Рис. 2.3.1

Длина саркомера составляет 2,5 – 3 мкм, диаметр – 1-2 мкм и выглядит как система чередующихся из дисков. Формула саркомера: Z+½ I + A +½ I + Z.
Он представляет собой систему тонких (thin, actin) актиновых и толстых (thick, myosin) филаментов
А - диск –зона локализации толстых миозиновых филаментов. Включает в свой состав несколько подзон:
М – линия (зона крепления миозиновых филаментов в центре саркомера)
Н – подзона (диск) -участок расположения основной массы миозиновых протофибрилл. В пределах этой зоны, исходя из положения одного из важнейших белков саркомера – титина,выделяют ряд участков: С – участок,где нет перекрытия с актиновыми нитямии D – участок,где миозиновые фибриллы истончаются и есть перекрытие с концами актиновых филаменто
I - подзона (диск) –участок расположения актиновых фибрилл вне зоны перекрытия с миозиновыми фибриллами. В каждом саркомере располагается с двух сторон ½ I диска.
Z - линия (диск) –граница соседних саркомеров.
Современные исследования структуры и химического состава саркомеров показали, что они имеют очень сложную пространственную организацию, а в его состав, помимо основных сократимых белков актина и миозина входит более двух десятков вспомогательных белков, принимающих участие в функционировании саркомера и мышцы в целом.
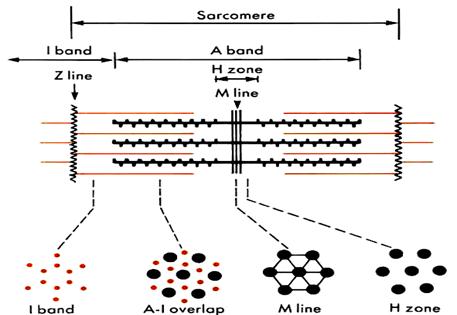
Рис. 2.3.2. Традиционная схема саркомера и его деления на диски

Актиновые и миозиновые протофибриллы образуют сложную упорядоченную сеть. У позвоночных животных встречается два типа «актин-миозиновой решетки», хотя соотношение актин/миозин всегда одно 3:1.
- «суперупорядоченная решетка», характерная для высших позвоночных (млекопитающих и птиц).
- «простая решетка»,характерная для саркомеров мышц низших позвоночных (рыб и амфибий)
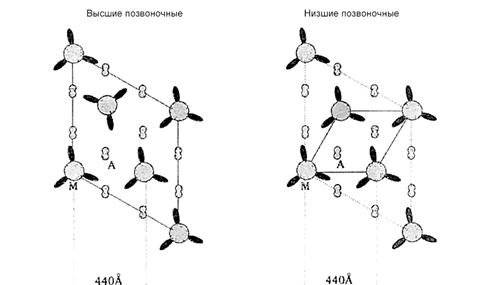
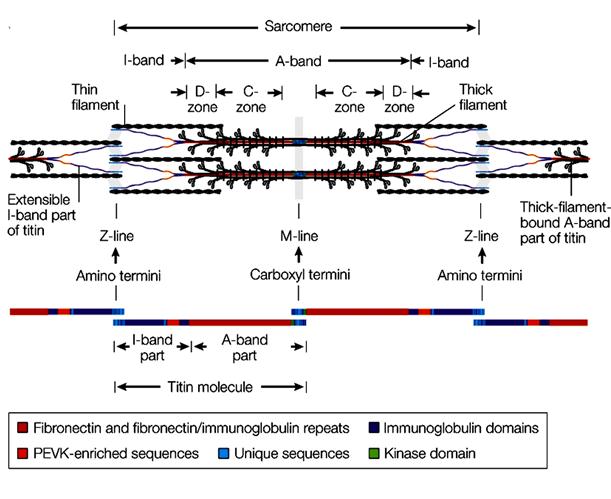
Рис. 2.3.4. Современная схема организации саркомера и расположения в нем белка титина ( по: L.Tskhovebova, J.Trinick, 2003; Physiol.Rev, 2005)
Саркоплазматический ретикулум – представляет собой специализированную сеть гладкого эндоплазматического ретикулума. (L – система). Канальцы СР идут вдоль миофибриллы, но канальцы соседних саркомеров изолированы друг от друга, тогда как СР соседних миофибрилл имеют систему соединительных трубочек. Продольные канальцы СР охватывают миофибриллу со всех сторон, образуя т.н.»воротничок».
По ходу мышечного волокна плазмалемма образует систему поперечных впячиваний – Т- каналов (Т-система). В районах Т-каналов канальцы саркоплазматического ретикулума сливаются, образуя т.н. «терминальные цистерны».Т-каналы могут находится как на уровне Z- дисков (рыбы, амфибии), так и по длине саркомера на уровне А и I –диска (круглоротые, рептилии, птицы и млекопитающие).
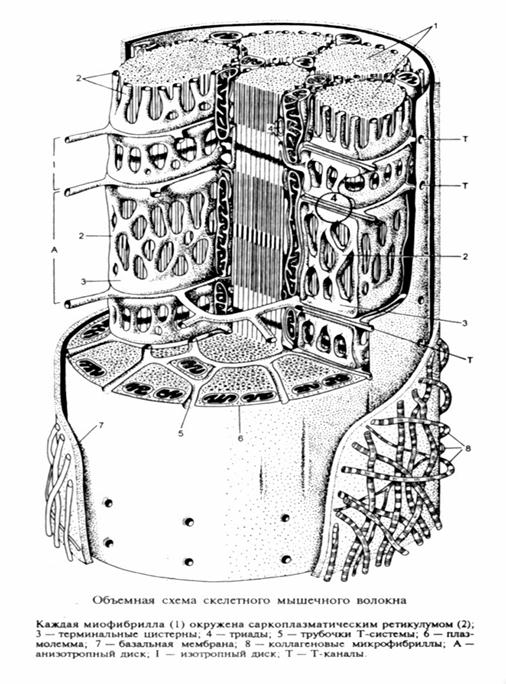
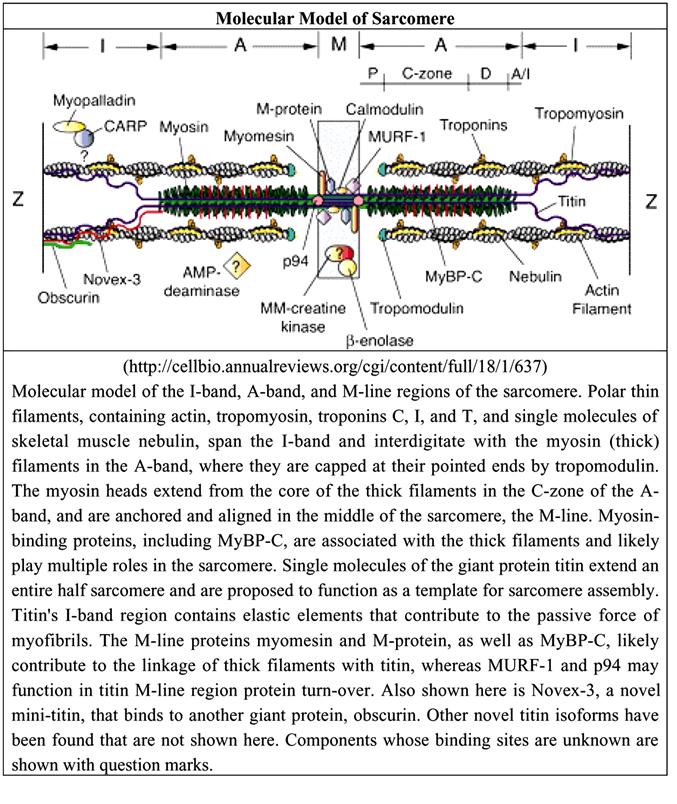
Рис. 2.3.5. Молекулярная структура саркомера (по:E.Niederhoffer, Muscle biochemistry, 2004).
Таким образом каждая трубочка Т-системы с двух сторон контактирует с терминальными цистернами L – системы в пределах каждого саркомера, образуя т.н. «триады».Система L – и Т- каналов значительно различается в тонических и фазных волокнах.
В фазных волокнах, как отмечалось выше, триады располагаются в каждом саркомере не только на уровне Z – дисков, тогда как в тонических мышцах, триады всегда располагаются только на уровне Z – дисков (Кроленко, 1975).
Митохондрии– мышечного волокна очень хорошо развиты, что связано со значительными энергетическими затратами при движении. В связи с этим, в мышцах встречается особый тип митохондрий – митохондриальный ретикулум.
Он представляет собой гигантские разветвленные митохондрии, оплетающие миофибриллы. Между этими пластами ретикулярных митохондрий расположены своеобразные мостики из т.н.нитчатых митохондрий. Тем самым образуется трехмерная митохондриальная сеть, части которой соединены специальными межмитохондриальными контактами (ММК). Через эти контакты возможна связь между отдельными, хотя и гигантскими ретикулярными и нитчатыми митохондриями, объединяя их в единую энергетическую систему мышечного волокна.
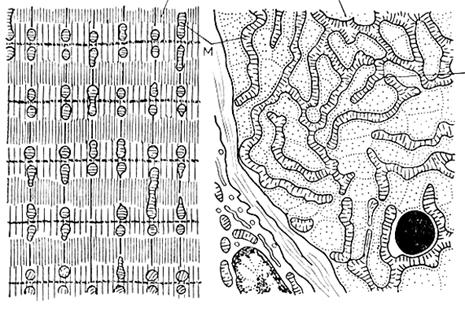
А – продольный срез, б – на уровне Z – диска (Ченцов, 1995).
Помимо гигантских ретикулярных митохондрий, в мышечных волокнах встречаются митохондрии традиционного типа – отдельные, мелкие митохондрию Особенно много их в околоядерной области.
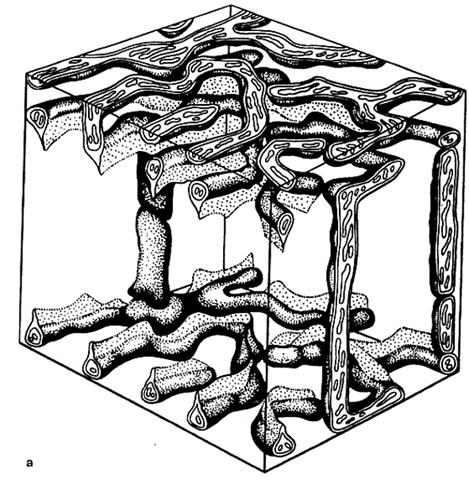
Рис. 2.3.7. Объемная реконструкция участка митохондриального ретикулума в мышечном волокна соматической (скелетной) мышце млекопитающих (крыса) (Бакеева, 1980, Шубникова и др., 2001).
Аппарат Гольджи развит не очень сильно, также как и рибосомальная система. Необходимо только отметить, что актин и миозин синтезируются на полисомах, включающих от 15 до 75 рибосом.
В саркоплазме мышечных волокно обнаружено большое количество специфического белкового пигмента – миоглобина.Именно его количество и определяет принадлежность волокон к «красному» или «белому» типам. Также встречаются скопления гликогена(особенно его много в белых мышечных волокнах), тогда как липидных капель – наоборот мало.
В настоящее время биохимическая организация мышечных волокон скелетной мускулатуры позвоночных (особенно млекопитающих) изучена достаточно полно.