Циркуляция биологического времени на уровне целого организма растения.
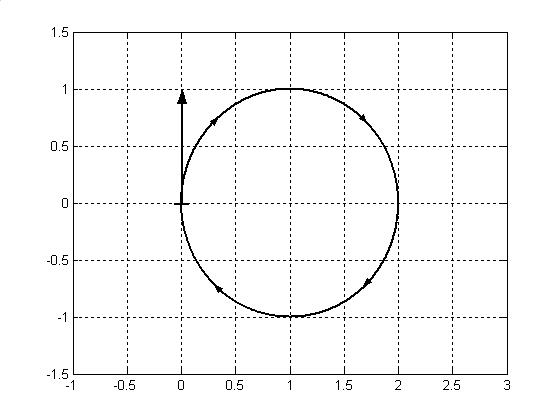
Замкнутый криволинейный интеграл в основе своей предполагает существование некоторой замкнутой кривой, по которой берется интегрирование. Замкнутая кривая при этом может иметь самый разнообразный вид. Эта кривая может быть как с самопересечениями, так и без. Кривая без самопересечений в наиболее простом виде всегда гомеоморфна окружности, как простейшей замкнутой кривой. Поэтому, в первом приближении, в качестве замкнутой кривой, по которой берется интегрирование, будем рассматривать окружность, рис.5. Такая окружность может существовать лишь в двумерном временном пространстве.
отн. ед. биол. времени
отн. ед. биол. времени
Рис.5. Движение биологического времени (циркуляция) по окружности в течении онтогенеза растения. Показан единичный вектор скорости движения как касательный вектор к окружности.
Теперь покажем, что данный характер движения биологического времени (циркуляция по окружности) соответствует существующим физиологическим представлениям о росте и развитии организма растения.
Будем считать, что вектор скорости движения биологического времени по окружности единичен. Теперь найдем проекции этого вектора скорости на первую (ось абсцисс) и вторую (ось ординат) временные оси. Первая проекция составляет величину:
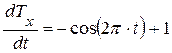 , (7)
, (7)
где 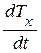 - проекция вектора скорости биологического времени Т на ось абсцисс, отн. ед. биол. времени; t – физическое время, выраженное в целых единицах времени всего онтогенеза. Переменная физического времени t изменяется в диапазоне 0 ≤ t ≤ 1, тогда точка движения по окружности пройдет весь оборот, соответствующий всему периоду онтогенеза.
- проекция вектора скорости биологического времени Т на ось абсцисс, отн. ед. биол. времени; t – физическое время, выраженное в целых единицах времени всего онтогенеза. Переменная физического времени t изменяется в диапазоне 0 ≤ t ≤ 1, тогда точка движения по окружности пройдет весь оборот, соответствующий всему периоду онтогенеза.
Вторая проекция (на ось ординат) находится из соотношения:
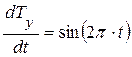 , (8)
, (8)
где 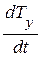 - проекция вектора скорости биологического времени Т на ось ординат, отн. ед. биол. времени.
- проекция вектора скорости биологического времени Т на ось ординат, отн. ед. биол. времени.
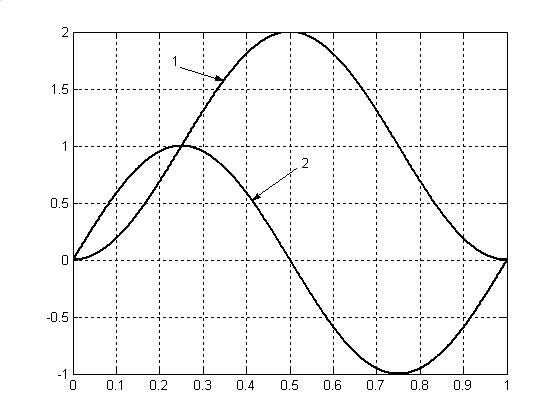
Полученные зависимости первой и второй проекции скорости движения биологического времени по окружности представлены на рис.6.
Как видно из полученных данных на рис.6, первая проекция вектора скорости циркуляции биологического времени представляет собой не что иное, как нормированную скорость накопления общей биомассы организма растения в онтогенезе в результате процесса фотосинтеза. При этом, первая проекция достаточно хорошо подтверждает экспериментальный закон «треугольника», предложенный А.А. Ничипоровичем для зерновых культур [7], Х..Г. Тоомингом для картофеля [6], а также подтвержден нами для подсолнечника [8]. Кроме того, сравнивая кривую 1 на рис.6 и кривую роста общей биомассы растения на рис.2 мы видим полную аналогию.
Вторая проекция впервые получена нами для двумерного временного пространства организма растения. Эта проекция характеризует динамику процесса общего дыхания растения в онтогенезе. Как видно из полученных данных на рис.6, вторая проекция, характеризующая интенсивность дыхания, в первую половину онтогенеза положительна. Это обеспечивает увеличение интенсивности дыхания до середины онтогенеза. Во вторую половину онтогенеза вторая проекция отрицательна. Это характеризует снижение интенсивности дыхания во вторую половину онтогенеза.
Следует отметить, что полученные численные характеристики процессов общего фотосинтеза и общего дыхания растения в онтогенезе впервые получены нами теоретическим путем на основе единого процесса –
отн. ед. биол. времени
отн. ед. биол. времени
Рис.6. Проекции скорости циркуляции биологического времени Т на ось абсцисс dTx/dt (1) и ось ординат dTy/dt (2).
циркуляции биологического времени в организме растения. Такой подход впервые позволяет объединить на единой основе процессы фотосинтеза и дыхания, а также позволяет проводить детальный анализ этих двух составляющих продукционного процесса растений и находить причины их проявления.
Показателен также интеграл уравнений (7) и (8). Интеграл первой проекции вектора скорости циркуляции биологического времени по окружности (7) , характеризующий процесс фотосинтеза, представлен уравнением:
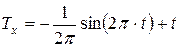 . (9)
. (9)
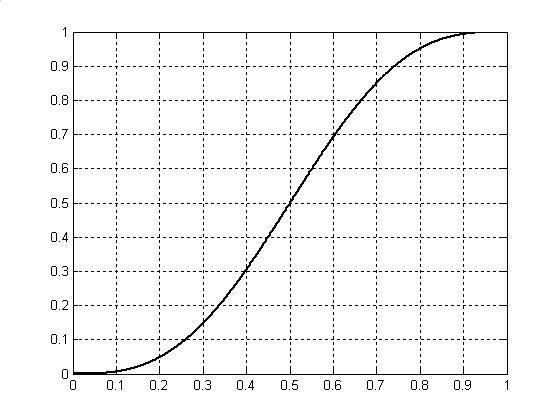
Графическое изображение интеграла (9) дано на рис.7. Как видно из рисунка 7, мы получили не что иное, как логистическую кривую роста общей биомассы организма растения μ в онтогенезе. Эта логистическая кривая получена теоретически и характеризует прохождение процесса фотосинтеза растения в онтогенезе. В тоже время, логистическая кривая характеризует изменения биологического времени в целом организме растения как проекция на временную ось «прошлое-будущее».
(отн. ед. биол. времени)2
отн. ед. биол. времени
Рис.7. Интеграл первой проекции вектора скорости биологического времени на ось абсцисс.
Интеграл второй проекции вектора скорости циркуляции биологического времени по окружности (8), характеризующий процесс дыхания, представлен уравнением:
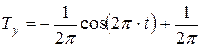 . (10)
. (10)
Как видно из полученного уравнения (10), временные процессы дыхания представлены двумя составляющими. Первая составляющая дыхания представляет собой динамический процесс через переменную физического времени t. Вторая составляющая является постоянной величиной. Таким образом, описанный временной процесс дыхания состоит из двух компонент: динамической и постоянной. Сравнивая уравнения (7) и (10), мы получаем:
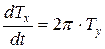 . (11)
. (11)
Из последнего уравнения (11) видно, что временные процессы дыхания включены во временные динамические процессы фотосинтеза, что является непосредственным теоретическим обоснованием экспериментальной гипотезы Мак-Кри о двухкомпонентном дыхании: фотодыхание и дыхание поддержания.
Выводы:
1. Метод сумм эффективных температур эффективен до фазы «цветение» растений. В тоже время этот метод позволяет ввести в рассмотрение двумерное временное пространство организма растения.
2. На примере незамкнутого криволинейного интеграла биологического времени растений показано, что существенными влияющими силами на организм растения является биологическое время. Рассматривается физическое и биологическое время, при этом биологическое время подвержено влиянию на него агрометеорологических условий вегетации.
3. На единой основе циркуляции биологического времени в организме растения объединены процессы фотосинтеза и дыхания растений. Показана роль процессов циркуляции биологического времени для процессов роста и развития.
4. Теоретически, на основе циркуляции биологического времени, подтверждена экспериментальная гипотеза Мак-Кри о двухкомпонентном дыхании растений.
5. Данный подход к проблеме биологического времени организма растения позволяет проводить глубокий анализ фундаментальных процессов онтогенеза, таких как фотосинтез, дыхание, рост, развитие и объединять эти процессы на единой основе.
Литература
1. Наумов М.М. Векторный характер биологического времени растений. // Метеорологія, кліматологія та гідрологія. -2004. –Вип.48. С.226-234.
2. Миусский П.Е., Наумов М.М., Русакова Т.И. О математической модели продукционного процесса подсолнечника.// Метеорология, климатология и гидрология. -1989. –Вып.24. С.132-137.
3. Наумов М.М. Прогноз урожайности семян подсолнечника на основе математической модели продукционного процесса.// Матеріали міжнародної конференції „Гідрометеорологія і охорона навколишнього середовища – 2002”. –Одеса. -2003. –Ч. 1. С.264-270.
4. Полевой А.Н. Теория и расчет продуктивности сельскохозяйственных культур. –Ленинград, Гидрометеоиздат, 1983, -175с.
5. Сиротенко О.Д. Математическое моделирование водно-теплового режима и продуктивности агроэкосистем. –Ленинград, Гидрометеоиздат, 1981, -167с.
6. Тооминг Х.Г. Экологические принципы максимальной продуктивности посевов. –Ленинград, Гидрометеоиздат, 1984, -264с.
7. Ничипорович А.А. Теория фотосинтетической продуктивности растений. В кн..: Итоги науки и техники, сер. Физиология растений. –Москва, ВИНИТИ, 1977, т.3. С.11-54.
8. Наумов М.М. Фотосинтетическая деятельность культуры подсолнечника в посеве.// Метеорологія, кліматологія та гідрологія. -2003. -Вип.47. С.118-125.