ОСНОВЫ БИОТЕХНОЛОГИИ
1. Основные направления биотехнологии.
2. Пути регуляции клеточного метаболизма.
3. Механизмы регуляции катаболизма клетки.
4. Механизмы регуляции анаболизма клетки.
5. Регуляция метаболизма в разветвленных схемах биосинтетических превращений.
6. Биосинтез первичных метаболитов.
7. Биосинтез вторичных метаболитов.
Биотехнология – это использование микроорганизмов, культур клеток животных и растений, метаболизм и биосинтетические способности которых обеспечивают выработку специфических, необходимых человеку веществ.
Возникновение и развитие биотехнологии базируется прежде всего на использовании микроорганизмов.
Основные направления биотехнологии:
1. Генная инженерия – это раздел биотехнологии, который изучает способы модификации ДНК и перенесение ее из одних организмов в другие. Возникла благодаря интенсивному прогрессу вирусологии, бактериологии, генетики, энзимологии, молекулярной биологии.
Цель: встраивание природной ДНК в плазмиду или геном, такая клетка дает клоны трансформирующих клеток, способных к образованию в больших количествах специфических целевых веществ. Цель – создание высокопродуктивных штаммов для традиционных микробиологических процессов. Достижения генетики используются в селекции, т.е. выведении новых форм организмов со свойствами, резко отличающимися от свойств исходных форм.
Методы селекции:
1). Естественный отбор
2). Искусственный отбор
3). Искусственный отбор измененных форм, индуцированных мутаенезом.
4). Получение измененных форм путем гибридизации, когда объединенные полезные признаки исходных форм активизируются в гибриде.
2. Микробная биотехнология.
3. Клеточная инженерия
4. Химическая энзимология (ферментология)
5. Инженерная энзимология
 С помощью биотехнологии получают антибиотики, витамины, ферменты, аминокислоты, нуклеотиды, стероиды, гормоны, интерфероны, вакцины, диагностические препараты, антитела, органические кислоты, биополимеры, биопестициды и т.д.
С помощью биотехнологии получают антибиотики, витамины, ферменты, аминокислоты, нуклеотиды, стероиды, гормоны, интерфероны, вакцины, диагностические препараты, антитела, органические кислоты, биополимеры, биопестициды и т.д.
2. Микроорганизмы осуществляют множество реакций, которые катализируются различными ферментами.
Все эти процессы катализируют сотни ферментов, которые в свою очередь синтезирует клетка
Регуляция клеточного метаболизма идет на двух уровнях:
1. На уровне регуляции синтеза ферментов.
2. На уровне регуляции активности ферментов
РЕГУЛЯЦИЯ НА УРОВНЕ СИНТЕЗА
Синтез каждого фермента регулируется отдельным геном. Несмотря на постоянство генотипа микроорганизмы очень подвижны в отношении к своему составу и метаболизму в ответ на окружающую среду.
Существуют два вида ферментов:
1. Ферменты, которые синтезируются вне зависимости от окружающей среды – это конститутивные ферменты (эндоферменты) или ферменты анаболизма (синтеза). Их образование осуществляется путем регуляции по принципу обратной связи, а именно ингибирования или репрессии.
2. Ферменты, которые образуются только тогда, когда в среде присутствуют их субстраты. Это так называемые индуцибельные ферменты или ферменты катаболизма, т.е. те ферменты, которые участвуют в использовании субстратов (питательных веществ) -
экзоферменты. Изменение количества индуцибельных ферментов в клетке происходит под действием индукции или катаболитной регуляции.
РЕГУЛЯЦИЯ НА УРОВНЕ АКТИВНОСТИ ФЕРМЕНТОВ
Регуляция на уровне активности ферментов свойственна ключевым ферментам обмена веществ. Активность повышается под действием положительного эффектора и снижается под действием ингибитора (отрицательного эффектора).
РЕГУЛЯЦИЯ СИНТЕЗА ФЕРМЕНТОВ КАТАБОЛИЗМА КЛЕТКИ
1. Индукция – это регуляция синтеза ферментов реакций разложения. Индукция относительно повышает скорость синтеза ферментов в ответ на присутствие в среде определенного химического вещества – индуктора (субстрат, его аналог, продукты реакции). Индукция обеспечивает клетке возможность использовать аминокислоты и энергию для синтеза тех ферментов, которые необходимы ей в данный момент.
Наиболее распространена модель Жакоба – Моно
 ДНК
ДНК
| R | P | O | S |
R – ген, закодированный на синтез активного белка – репрессора.
Р – ячейка, где находится РНК – полимераза
О – ген – оператор, управляет работой структорного гена S
S – структурный ген, ответственный за синтез целевого белка
Если в среде нет индуктора, белок – репрессор соединяется с геном и блокирует прохождение РНК – полимеразы к гену S. Таким образом, РНК – полимераза, катализирующая транскрипцию ДНК в информационную РНК, не достигает гена S, следовательно не проходит синтеза белка, или трансформации. Если в среде присутствует индуктор, он связывается с белком – репрессором, изменяет его структуру, переводя а неактивное состояние. Белок – репрессор в таком состоянии не может связаться с геном О. РНК – полимераза проходит вдоль молекулы ДНК через ген О, который включает ген S. Происходит транскрипция, а затем трансляция.
2. Катаболитная регуляция – если в среде находится несколько субстратов, которые клетка способна использовать, она выбирает наилучший и вырабатывает ферменты для его использования. Обычно это глюкоза. По мере ее потребления синтезируется фермент для расщепления другого субстрата и т.д.
РЕГУЛЯЦИЯ СИНТЕЗА ФЕРМЕНТОВ АНАБОЛИЗМА
4.1. Репрессия: ген R закодирован на синтез неактивного белка апорепрессора. Если в среде присутствует корепрессор (продукт реакции), он взаимодействует с апорепрессором и переводит его в активное состояние. Корепрессор и апорепрессор взаимодействуют с геном О и блокируют движение РНК – полимеразы. Синтеза белка не происходит. Если корепрессор отсутствует, неактивный апорепрессор не может соединиться с геном О. РНК- полимераза проходит к гену S, который включается в работу.
2. Ингибирование: в этом случае ингибитором может быть конечный продукт реакции. Каждый фермент имеет два центра: активный – для связывания субстрата и ингибиторный – для связывания ингибитора.

Если ингибитор соединяется с ферментом, то изменяется структура активного центра, что затрудняет взаимодействие с субстратом. Этот эффект называется аллостерическим. Зависимость скорости реакции от концентрации субстрата имеет следующий вид

Регуляция синтеза ферментов анаболизма называется регуляцией по принципу обратной связи.
5. Если клеткой в общей цепи метаболических реакций
F
 f
f
а в с d
 А →В →С →Е →D
А →В →С →Е →D
k
К
образуется не один, а несколько продуктов (А,В,С,Е,К,D,F) с участием соответствующих ферментов (a,в,c,d,f,k), то существует ряд механизмов регуляции синтеза этих метаболитов.
Первый механизм: дифференцированная регуляция изоферментами. Допустим, что А→В – реакция превращения А в В катализируется несколькими изоферментами, но синтез каждого из них контролируется различными конечными продуктами.
Второй механизм: согласованная регуляция по принципу обратной связи. Превращение вещества А в В катализируется одним ферментом, но образуется несколько конечных продуктов, которые согласовано ингибируют его синтез.
Третий механизм: кумулятивная регуляция по принципу обратной связи, каждый конечный продукт способен вызвать очень слабое ингибирование/репрессию синтеза фермента, но комбинация конечных отдельных продуктов имеет кумулятивный характер.
6. Биосинтез первичных метаболитов. Первичные метаболиты – это низкомолекулярные соединения необходимые для роста микроорганизмов. Их молекулярная масса менее 1500 дальтон. Наиболее важные среди них - аминокислоты, органические кислоты, пуриновые и пиримидиновые основания. Микробная клетка не производит избытка этих метаболитов, а следовательно не накапливает их, но есть микроорганизмы с нарушением обмена веществ, их используют в биотехнологии для получения необходимых веществ.
Разработано две методики получения первичных метаболитов:
1. Накопление промежуточных продуктов, или ограничение синтеза конечного продукта.
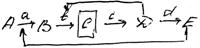
Существует цепь метаболических реакций, каждый из этих этапов катализируется своим ферментом. Нам необходимо получить промежуточный продукт С. Продукт Е подавляет синтез фермента а, D – подавляет синтез в. Чтобы накопить вещество С, получают мутантный штамм, у которого отсутствует фермент с.

Если к питательной среде (клетке) подводится недостаточное количество вещества Е или D, то репрессии синтеза ферментов а и в не происходит (сама клетка веществ Е и D в отсутствии фермента с не синтезирует) .
2. Накопление конечных продуктов рассматривается на примере синтеза аминокислоты – лизина; бактериями лизин синтезируется по разветвленной схеме.
Исходное вещество аспартат, который под действием фермента Ф1 превращается в аспартилфосфат, а затем в ряд аминокислот: лизин, треонин, лейцин, метионин.
аспартат
↓Ф1
аспартилфосфат
↓
аспартилполуальдегид

 Ф2
Ф2

 лизин гомосерин
лизин гомосерин
 |
треонин метионин
лейцин
Без фермента Ф2 – только в лизин, а с ферментом Ф2 – в треонин, лейцин и метионин.
У каждой из этих аминокислот механизм согласованной регуляции на синтез фермента Ф1.Если используются микроорганизмы, у которых отсутствует фермент Ф2 ,то ингибирующий эффект на синтез фермента Ф1 будет недостаточным. Лизин будет синтезироваться в больших количествах.
7. Синтез вторичных метаболитов – это молекулы веществ, которые синтезируются на последних стадиях ростового цикла, они не требуются для роста организма, но играют существенную роль в его жизнедеятельности.
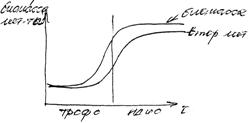
Весь цикл роста и развития организма можно разделить на две фазы:
1. Трофофаза – фаза начального роста.
2. Идиофаза – фаза конечного роста.
Ко вторичным метаболитам относятся: антибиотики, гормоны, алколоиды, токсины.
При биосинтезе вторичных метаболитов установлено влияние следующих регуляционных механизмов:
1. Индукция ферментов: индуктор стимулирует синтез фермента, но в структуру вторичного метаболита не включается.
2. Регуляция по принципу обратной связи (репрессия конечным продуктом в разветвленной схеме биосинтеза).
3. Катоболитная регуляция (используется для получения пенициллина).
При выращивании гриба Penicillium notatum – продуцента пенициллина в питательную среду вносят смесь глюкозы и лактозы. Глюкоза необходима для роста, лактоза расходуется медленно, но хорошо поддерживает синтез пенициллина. Поэтому целесообразно в трофофазе добалять глюкозу, а в идиофазе – добавлять лактозу.
8. БИОТРАНСФОРМАЦИЯ, ФАКТОРЫ ЕЕ ОПРЕДЕЛЯЮЩИЕ.
Биотрансформация – это неполное превращение органических соединений ферментами микроорганизмов. Используются растущие клетки для получения веществ, которые невозможно получить другим путем. При трансформации участвуют один или несколько ферментов, поэтому исходное вещество меняется незначительно. Трансформация используется для получения веществ, которые невозможно получить другим путем, например, из глюкозы получают глюконовую кислоту. Д-фенилаланин превращается в L-фенилаланин.
Преимущества в сравнении с химическим синтезом:
1. Специфическое действие микробных ферментов позволяет осуществлять тонкую перестройку молекул органических веществ с помощью простых технических схем, минуя многостадийные синтезы.
2. Ферменты действуют в условиях, не требующих повышенной температуры.
3. Образуется небольшое количество побочных продуктов, в т.ч. засоряющих окружающую среду.
4. Получают большой выход биомассы.
Факторы, определяющие биотрансформацию:
1. Регуляция синтеза ферментов, для которой используют индукцию. В среду добавляют индуктор -субстрат или его аналог. В этом случае предотвращается катаболитное подавление синтеза нужных ферментов.
2. Используют мутантные штаммы, у которых отсутствует фермент, разлагающий целевой продукт.
3. Используют смесь двух субстратов, из которых один потребляется микроорганизмом для роста, а другой превращается в целевой продукт.
4. Используется проницаемость микробных мембран, то есть осуществляется регуляция поступления веществ за счет синтеза определенных пермеаз.
9. РЕГУЛЯЦИЯ МИКРОБНОГО СИНТЕЗА ФЕРМЕНТОВ.
Для получения в биотехнологии целевого фермента осуществляют:
1. Выбор продуцента: проводится скрининг штаммов и выбирают наиболее активный по способности синтезировать нужный фермент.
2. Разрабатываются условия выращивания этого микроорганизма с целью максимального биосинтеза фермента: подбор питательной среды, определенная температуры для роста, рН, уровень аэрации.
3. Изучается динамика биосинтеза, т.е. определяется время выращивания, соответствующее максимальному накоплению фермента.
4. Разработка схемы выделения и очистки фермента.
В процессе культивирования используется механизм индукции. Чтобы исключить катоболитное подавление синтеза, из питательных сред выводят легкодоступные источники углерода. Чтобы подавить репрессию, ингибирование, т.е. синтез веществ, действующих на синтез нужного фермента, из среды выводят продукты синтеза.
Преимущества микробного синтеза ферментов:
1. В качестве питательной среды используют дешевое сырье (отходы производств).
2. Цикл ферментации достаточно короткий: 36-40 часов при поверхностном культивировании, 48- 70 часов – при глубинном.
3. Простые процессы фракционирования и разделения ферментов.
4. Микроорганизмы способны к сверхсинтезу.