Лекция 1. Введение.
Это аморфные и микро- скрыто- или полукристаллические минералы. Хотя среди них и встречаются высококристаллические (даже гигантокристаллические ) минералы. Это глинистые, кремневые, железистые, Al, Mn, органические.
Общие свойства аутигенных минералов
2. Гидратированность, причём вода в их составе разная:
- гидроксильная (входит в решетку),
- цеолитная ( в каналы решетки),
- сорбционная (гигроскопическая),
- кристаллогидратная (образует твердый раствор).
3. Они имеют переменный состав: не только переменный состав воды, но и изменчивый комплекс поглощённых катионов и других компонентов (адсорбирование редких и рассеянных элементов). В них широко распространены изоморфизм, полиморфизм, что осложняет выделение минеральных видов, также осадочные минералы чаще магматических представлены в окисленной форме. Всё это затрудняет изучение осадочных пород.
4. Способность реагировать на изменение окружающей среды (как у животных). Особенно сильно это проявляется в самых типичных и распространённых глинистых минералах (50 % от всех осадочных минералов) с подвижной кристаллической решёткой, способной раздвигаться и сжиматься в зависимости от увеличения или уменьшения влажности, обладающей возможностью катионного обмена с другими минералами и средой. Ёмкость поглощения у глин наибольшая из всех минералов и она разнообразна по природе: катионы и биполярные молекулы воды входят между слоями кристаллической решётки, поглощаются огромной поверхностью глинистых частиц, а также чисто капиллярными силами.
Отличия глауконита аллотигенного от аутигенного
Аутигенный:
1) корочки железа,
2) крупная поляризация,
3) форма угловатая и изрезанная.
Аллотигенный:
1) нет корочек железа - содраны,
2) мелкая поляризация,
3) форма окатанная.
Литологический анализ правильнее начинать не с отдельных слоёв, а с их парагенезов, выделением многослоёв (циклитов). Парагенезы слоёв раскрывают строение толщ или формаций. Осадочный слой всегда обнаруживает ту или иную цикличность, т.е. свойство распадаться на комплексы тесно взаимосвязанных слоёв, отвечающих законченным фазам или этапам развития той или иной области седиментации. Особое внимание обращается на взаимопереходы пород внутри слоя. Фиксируется мощность слоёв, их вертикальная анизотропия, характер смены (постепенный, плавный), смену включений и других признаков, горизонтальную изменчивость, кровлю и подошву.
Естественно, выявляются признаки свидетельствующие о механизме и условиях образования и даётся первый вариант их генетического истолкования, выражающийся в отнесении слоёв к тем или иным генотипам.
Nota bene Нота бене - обратить внимание
Некодирующие РНК не участвуют в кодировании белков, но составляют очень значительную часть общего транскриптома. Огромная роль в регуляции генов на уровне транскрипции, на уровне пост-транскрипционного процессинга и на уровне трансляции. Эпигеномика – наложение «нестабильной информации» на информацию, которая кодирует нуклеотидная последовательность. Внешние воздействия могут влиять на этот слой информации, это называется – эпигенетическая изменчивость (находится «над генетическими детерминантами, закодированными в последовательности ДНК»).
В университете дружбы народов цветет «лысенконизм»))
Вы видите знаменитый ландшафт Уоддингтона (он предложил термин «эпигенетика», работал с развитием дрозофилы и исследовал достаточно нестабильные (обратимые) наследуемые изменения, которые назвал эпигенетическими), сейчас один из самых цитируемых. Ландшафт показывает, что есть клетка, которая при дифференцировке может попасть в одну из долин этого ландшафта (это разные состояния этой клетки). На вершине хребта – нестабильное состояние, с которого она скатывается в зависимости от воздействий на эту клетку. Чем выше хребет, тем глубже долина, тем более стабильное состояние. Канализация (этот термин тоже ввел Уоддингтон) – будем рассматривать при рассмотрении микроРНК – идет развитие по определенному каналу и никуда не может уйти. Уоддингтон ввел «эпигенетику» - в 40-50гг, Уотсон как раз открыл двойную спираль (2 статьи в Nature, так как с первого раза их не поняли), Уоддингтон не понял, когда ему рассказывали (был не очень восприимчивый, бедняга).

Очень небольшая часть генома (жалкие проценты, около 2-4) участвует в кодировании белков, основная масса ДНК не участвует в кодировании белков, но она все равно транскрибируется. Более 90% генома транскрибируется, в том числе гетерохроматин (транскрипция этого участка показывает, что затем этот участок генома будет неактивный, есть определенные эпигенетические механизмы для этого, которые в том числе наследуются в клеточных поколениях). Геном – машина, которая синтезирует РНК, и в основном это некодирующая РНК (в том числе рибосомальная), которая осуществляет регуляторную роль, несвязанную с кодированием белков. Молекулы мРНК содержат определенные участки, достаточно длинные, которые не участвуют в кодировании белков, а нужны для регуляции трансляции этой РНК, но не только для этого нужна эта некодирующая.
Сейчас вспоминают статью 1961 Жакоба и Моно – определенная модель регуляции экспресии генов с участием белков - но в начале они высказали мысль, что репрессором может быть молекула РНК, которая может найти комплементарную последовательность ДНК и участвовать в регуляции ее экспрессии.
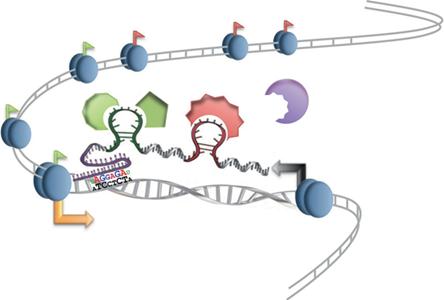
Сегодня нам покажут две вещи: 1) регуляторные нкРНК могут взаимодействовать с белками и другими РНК и вызывать их конформационные изменения, 2) и еще, что сам акт транскрипции меняет нуклеосомную структуру, и тем самым осуществляется динамика структуры хроматина (за счет транскрипции). Еще когда осуществляется транскрипция, новооброзованный транскрипт (еще висит на ДНК) – его узнают белки, модифицирующие белки хроматина и ДНК, и привлекаются к тому месту, где осуществляется транскрицпия (то есть не только определенные белки узнают определенные домены, но и просто транскрипты могут привлекать). Есть РНК со смешанными функциями: она может быть некодирующей, а в определенном случае может участвовать в кодировании белка.
(хвостик – CTD). Белок привлекается за счет транскрипции (образования определенной структуры):
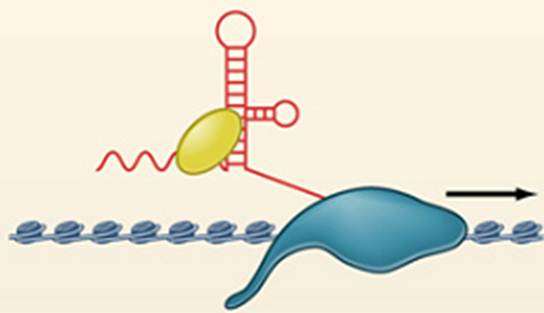
Структура РНК чрезвычайно динамична:

В РНК выделяется два самых главных компонента: стволик (за счет комплементарных взаимодействий) и петля. Дальше эта структура может перестраиваться таким образом, например, что 3’ (был комплементарный) отлипает и выступает как неспаренный участок, два U также неспарены, как и были. И нужно отметить, что когда функционируют нкРНК, им ассистируют РНК-шапероны (в данном случае РНК-хеликазы, которые расплетают двунитевые участки и меняют конформационную структуру РНК). Они могут не только расплетать, но и перестраивать рибонуклеопротеидный комплекс. Все нкРНК, о которых будем говорить, в комплексе с белками, поэтому это очень важная активность. Исследование нкРНК отстают в исследовании структур нкРНК, и сейчас многие исследователи этим занимаются.
Рибопереключатели: трансрикпция с помощью РНК-полимеразы, далее образуется транскрипт, который потом кодирует белок-насос, откачивающий избыток аденина у бактерий. Образуется шпилька, если нет аденина, – сигнал терминации – нет трансрикпции, белок-насос не образуется. Аденин меняет конформацию участка и шпилька образоваться не может. Транскрипция идет, насос образуется.
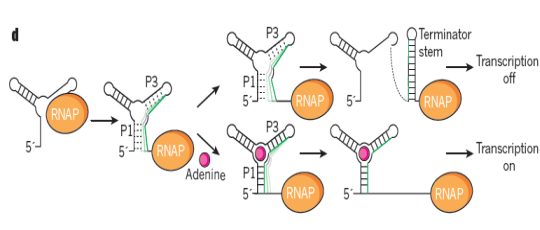
Что-то такое есть и в нкРНК.
Теперь общие слова о модификации хроматина с участием нкРНК. Нуклеосомная структура – елка с хвостами, на ней игрушки – химические модификации – гистоновый код. Белки, привлекающиеся с помощью транскрипции: есть читатели, есть писатели (на хвосты навешивают метки), все это работает согласованно и последовательно, еще есть комплексы, которые снимают метку. Могут создаваться негативные метки или позитивные метки, которые могут потом стираться. Модификации, которые эпигенетические (эпигенетическая метка – это не любая метка хроматина, а наследуемая).
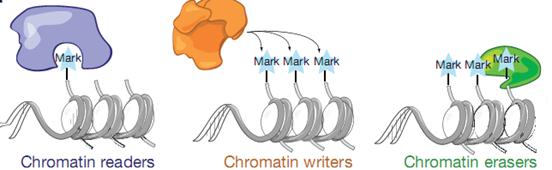
Скажем, (лизин – К) 27 лизин от конца гистона третьего триметилирован - в большей части репрсессорная метка, но иногда она может быть сильно активирующей, это зависит от контекста.
нкРНК привлекают белки к тому месту, где они образовались – цис-действие. Но не только. Есть человеческая нкРНК, которая образуется в одной части генома, а потом идет на другую хромосому:

На картинке – нкРНК с двумя доменами и она привлекает своими специфическими последовательностями (один домен) белок-писатель (негативная метка). А другой домен этой нкРНК привлекает белок, который стирает позитивную метку. То есть они работают согласованно.
Общий список:
• Число некодирующих транскриптов значительно больше, чем число кодирующих
• Сложная сеть перекрывающихся транскриптов, которые показывают, что 90% генома человека транскрибируется
• Относительное количество некодирующих регуляорных РНК увеличивается со сложностью организма (при усложнении организма меняются не только белки, но и способы регуляции)
• Транскрипция межгенных промежутков (раньше называли генными пустынями», а это матрица для нкРНК), энхансеров, промоторов и гетерохроматина
• Интроны (95% ДНК гена, экзон -150 н, интрон - 3500 н) – очень большая часть транскриптов.
Сначала думали, что такая повсеместная транскрипция – «транскрипционный шум», сейчас показана роль многих нкРНК, только механизмы неизвестны.
• Ассоциация полиморфизма ДНК в районах, не кодирующих белок, с болезнями человека. Оказалось, что это те районы, с которых образуется нкРНК.
• Консервативность (ультраконсервативность) в эволюции участков генома, транскрибируемых c образованием некодирующих РНК.
• Регуляция в развитии и тканеспецифичности.
Еще раз, нкРНК привлекают комплексы, модифицирующие хроматин. нкРНК и хромосомы ориентируют, и эти комплексы.
Еще дополнение к эпигенетическому ландшафту:

Есть эпигенетический барьер. Есть нуклеосомная структура. Хвосты гистонов могут быть ацетилированы и метилированы. И чтобы из метилированного состояния перейти в деметилированное и всякое подобное – нужна активация/инактивация соответствующих ферментов. Это перескок из одной долины в другую долину.
Слайд 14 не комментирует.
Эпигенетические метки:
1) ДНК-модификации
2) Гистоны, их модификации
3) Варианты гистонов (другой ген)
4) Негистоновые белки хроматина (осуществляют модификации)
5) РНК (очень существенная роль)
Классы нкРНК:
1. housekeeping (рРНК, тРНК, малые ядерные (в сплайинге) и ядрышковые РНК), давно известны
2. Короткие 20-35 н регуляторные РНК, чрезвычайно интересный класс
1. siРНК, small interfering (RNA interference) (без них никак изучать функции длинных нкРНК)
2. микро РНК
3. piРНК, связываемые белками PIWI (самый большой/обильный класс) (определяет иммунную системы и участвует в массе процессов)
3. Длинные (сотни-тысячи н), более 200 н (условно так считают)
lnc RNA, long non coding / linc RNA, long intergenic nc (linc - в межгенных пустынях)
4. Другие (ассоциированные с промотором, антисмысловые … )
Это картина распределения транскриптов. Видим интроны, экзоны. Типичная структура гена – основная масса ДНК – интроны. Около гена транскрипты и в противоположную сторону. Есть транскрипция, связанная с промотором (немного позже старта транскрипции). Очень сложный рисунок трансрипции, куча всего.

Может быть целое облако РНК около начала транскрипции мРНК (промотора).
Это удалось обнаружить благодаря новым технологиям детекции транскриптов – секвенирование, а также новым подходам, среди которых уничтожение образующихся транскриптов. Транскрипты не вечны, происходит смена одних на другие. Есть машина, которая уничтожает транскрипты. Если испортить ее мутациями, то можно увидеть больше транскриптов, в том числе с тех участков, где их немного или где они быстро уничтожаются. Наблюдается целый ряд нестабильных транскриптов, в обе стороны. В том числе перед промотором (тоже в обе стороны). У них куча названий. Есть транскрипты от конца гена, совсем непонятно что это такое – транскрипты, ассоциированные с терминатором транскрипции. Такое облако катологизируется. В общем, много активностей РНК-полимеразы 2 в обоих направлениях.

Итак, экзосомный комплекс уничтожает транскрипты (есть другая экзосома: клетка может секретировать пузырьки, которые внутри себя содержат различные биологически-активные материалы, среди них молекулы микроРНК. То есть экзосома это этот самый пузырек, она имеет мембрану (мембрана ЭПР), а внутри всякие штуки типа микроРНК. Она циркулирует в крови и может взаимодействовать с другими тканями).
А наша экзосома – для регуляции образованных транскриптов, а также для удаления неправильно-синтезированных транскриптов. Она видит, что РНК сложилась не как надо и ее уничтожает – наблюдение за точностью транскрипции. Совсем недавно появились сведения, которые говорят, что новообразованные транскрипты привлекают модификаторы гистонов (эпигенетические метки). Но и процессинг транскриптов с участием экзосомы тоже привлекает определенные модификаторы. В том числе в том месте, где идет транскрипция, затем идет процессинг (с помощью экзосом), это привлекает комплекс белков, которые делают его гетерохроматином.
Экзосома - контроль потока генетической информации, конкуренция с машиной процессинга (в каком то плане да, конкуренция, ведь если процессинг неверный, то экзосома ест транскрипт, или если не нужен процессинг, тоже ест). Экзосома осуществляет proof-reading транскриптов в клетках дикого типа (это для РНК-полимеразы, ведь она тоже ошибается). Экзосома разрушает СUT (сryptic untranslated transcripts) транскрипты (для чего они нужны неясно). Она обладает 3`- экзонуклеазной и эндонуклеазной активностями.
Что она из себя представляет. Она очень эволюционно-консервативна. Здесь перечислены ее разные субъединицы, которые очень схожи у дрожжей и человека, то есть очень консервативная и древняя.

То есть этот механизм регуляции давно возник и сохраняется. Ее основа – 9 субъединиц, которые составляют «шайбочку» (6 суб), на которую надевается «шапочка» (3 суб), косплекс не обладает нуклеазной активностью (для этого нужны еще другие субъединицы). В шапочку затягивается кончик РНК.
Кроме того, субъединицы, которые кушают РНК, садятся на эту основу (у них экзонуклеазная активность). А еще другая субъединица, с комплексом белков. В ней может быть и экзо- и эндо-нуклеазная активность. Видно, что все это очень похоже у людей и дрожжей.

Экзосомы могут быть в ядрышке, в нуклеоплазме, в цитоплазме. Все похожи. Но могут отличаться отдельными субъединицами.
Хвост, который затягивается, может поедаться. Путь этого транскрипта может быть и сразу в субъединицу (а может протягиваться через всю структуру).
Итак, несколько путей попадания РНК в экзосому (через шапочку или отдельно сразу туда, где поедается).

Есть некий комплекс, в который входит РНК-хеликаза, еще фиолетовая субъединица – полиА-полимеразная активность, еще другие белки. Этот комплекс узнает новообразованный транскрипт, осуществляет его полиаденилирование (но необычное, очень коротенькое полиА). А потом, когда этот комплекс соединился мишенью, то привлекается экзосома и начинается затягивание в нее конца РНК. Этот комплекс как раз и узнает неправильные конформации РНК. Чтобы экзосома функционировала, ей нужен однонитевой участок, который делает РНК-хеликаза.

Все это нужно для создания гетерохроматина.
Некодирующие транскрипты образуются тоже РНК-полимеразой II, на конце 5`-cap. Для них характерен типичный ландшафт хроматина - H3K4me3, H3K36me3 (позитивные метки). Процессинг осуществляется нормально (5`- cap, возможно полиаденилирование, сплайсинг). Многие некодирующие транскрипты подвергаются альтернативному сплайсингу (более 30% транскриптов), то есть это еще дополнение к регуляции. нкРНК может служить регуляторным сигналом, взаимодействуя с белками, РНК и ДНК. Может осуществлять комплементарные взаимодействия с РНК и непосредственно с ДНК. Дальше оказывается, что нкРНК везде присутствуют в клетке, есть типичные только ядерные, без полиА нкРНК (в специальных ядерных компартментах, обслуживают только транскрипцию, некий скелет для полимеризации внутриядерных частиц), есть ядернo-цитоплазматическая нкРНК, есть только цитоплазматическая. А есть вообще те, которые гуляют по организму.
Регуляторная роль нкРНК у про и эукариот осуществляется только в результате их непосредственного взаимодействия с белками. Есть определенные структурные элементы РНК, которые узнаются соответствующими белковыми доменами.
РНК-связывающие белки имеют разные домены, узнающие разные РНК-мишени, обладающие линейными специфичными нуклеотидными последовательностями или структурными мотивами. Такие белки иногда имеют домены, которые взаимодействуют и с РНК, и с ДНК. Итак, домены:
Ds RBD, double strand RNA binding domain (связывание двунитевых РНК)
RRM, RNA recognition motif, или RRD
Zn-finger, узнавание петель РНК (и для ДНК), очень динамичны
Гомеодомен (гомео-белки определяют развитие тела) (тоже и для ДНК)
Типичный домен – RRM. Интересны Arg/Lys-остатки, которые обеспечивают электростатические взаимодействия с фосфатами. Также часто наблюдаются стекинг-взаимодействия оснований РНК с ароматическими остатками.

А такой домен узнает с одного бока двуцепочечную РНК, он включает 70-90 аминокислот. Он узнает двунитевую РНК. Район 1 и 2 взаимодействует с малой бороздкой, район 3 - с большой. Домен узнает те участки, где находятся 2`OH и фосфаты, что дискриминирует дцДНК и РНК-ДНК (не узнает ДНК, только РНК). Показано, что узнает участок с одного бока, и это узнавание не сиквенс-специфично. Просто узнает определенную двунитевую структуру РНК.

Некодирующие малые РНК у бактерий, в том числе small RNAs (sRNA).
Этот класс – очень древний уровень регуляции. Важны у бактерий. Они вмешиваются практически во все физиологические функции, осуществляемые бактериями. Комплементарыне взаимодействия (которые в основе всего) схожие и у эукариот. Размер их около 100 нуклеотидов. Рассмотрим следующую картинку:

Левая стрелка - есть некий белок CsrA, который связывается с простенькой структурой: стволик+петля, которая на молекуле мРНК. Этот белок может осуществлять регуляцию трансляции соответствующей мРНК, укрепляя такие структуры. Дальше оказывается, что есть другая РНК (CsrB), которая имеет такую структуру, что каждый элемент этой структуры имитирует структуру мРНК, узнаваемую этим белком (стволик+петля). Эти белки переманиваются CsrB (РНК) и осуществляется регуляция трансляции.
Другая стрелка (средняя): есть 6S РНК (не достигает 200 нуклеотидов), которая имитирует структуру промотора. Промотор в обычных условиях открывается и образуется транскрипционный пузырь. А в плохих условиях как раз нужна такая 6S РНК, чтобы не было энергетических затрат на ненужную транскрипцию, на нее садится РНК-полимераза как на промотор и РНК-полимераза удаляется из пула транскрипции.
Правая стрелка - конкуренция между малыми РНК. РНК может быть дестабилизирована с участием соответствующего белка. Появляется другая РНК, которая этот белок забирает на себя и та РНК уже не дестабилизирована.
Дальше 1 слайд пропустил.
Ген у бактерия тоже открытая рамка трансляции и все дела. Есть транскрипция в противоположном направлении. Образуется антисмысловая РНК. С помощью комплементарных взаимодействий она соединяется и нет трансляции, это приводит к деградации. Это так называемое цис-действие. Где образовалась, там и действует.
Может быть транс-действие. Образуется в одном месте, а потом работает в другом месте. Ген далеко от места действия дает РНК, которая комплементарна ДНК в месте работы – некое транс-действие. Нет трансляции.
Может через расплетение шпильки хеликазами еще (расплетение закрепляется комплементарными участками).

А теперь перейдем к эукариотам.
Итак, мы рассматриваем два главных действия нкРНК у экариот.
1. воздействие молекулы нкРНК (взаимодействия с белками и комплементарные взаимодействия, вызывающие аллостерические эффекты)
2. роль акта транскрипции нкРНК, модифицирующего хроматин
Отдельные примеры этого.
Интересный случай – естественные антисмысловые транскрипты. И для полимеразы 2 и для полимеразы 1 (интез рРНК). Синтез рРНК – регуляция транскрипции с участием нкРНК. У арабидопсиса – для синтеза рРНК существует нормальный транскрипт. определяющий структуру рРНК и существует антисмысловой транскрипит. И чтобы заставить замолчать какую-то структуру ядрышка индуцируется синтез антисмыслового транскрипта, который приводит к понижению интенсивности смыслового транскрипта Или у дрожжей – тоже транскрипты в двух направлениях. И если антисмысловой транскрипт достаточно сильно производится, то это приводит в подавлению экспресии (в частности, фосфатазы 4). А другая фосфатаза – наоборот антисмысловой транскрипт привлекает что-то такое, что активирует транскрипцию этой самой фосфатазы.

Гипотетические представления об естественных антисмысловых транскриптах. Смысловой транскрипт осуществляет образование мРНК. Антисмысловой – привлекает своими структурами модификаторы (зелененький и красненький), которые записывают определенные метки. Кроме того, антисмысловой транскрипт приводит к образованию триплекса (между РНК и ДНК),что мешает смысловому транскрипту.