Темновая фаза фотосинтеза. Экология фотосинтеза.
План
1. Цикл Кальвина
2. Путь Хетча и Слэка
3. Метаболизм по типу толстянковых
4. Фотодыхание и метаболизм гликолевой кислоты
5. Эндогенные механизмы регуляции фотосинтеза
6. Влияние внешних факторов на процессы фотосинтеза
1. Цикл Кальвина (С3-путь фотосинтеза)
В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NADPH. Эти конечные продукты световой фазы фотосинтеза необходимы для темновой фазы, где СО2 восстанавливается до углевода. Темновая фаза фотосинтеза – сложный, многокомпонентный процесс. В настоящее время известно несколько путей метаболизма углекислого газа - С3 –путь, С4- путь, фотосинтез по типу толстянковых (САМ – метаболизм) и фотодыхание.
С3-путь фотосинтеза присущ всем растениям. Он был расшифрован 1946-1956 гг. американским биохимиком М.Кальвином и сотрудниками. Данное открытие было возможным благодаря использованию новейших ( на тот период времени) методов – двумерной хроматографии и авторадиографии (использование меченных атомов).
Для выявления первичного акцептора СО2 были использованы одноклеточные зеленые водоросли (хлорелла и сценедесмус) и меченный 14СО2. Водоросли помещали в среду, содержащюю 14СО2, на разные промежутки времени, затем клетки быстро фиксировали, экстрагировали и определяли содержание 14С в различных соединениях после их разделения с помощью хроматографии. Выяснилось, что после экспозиции в течение 1 мин меченный углерод включался в С3-С7-сахара, фосфосахара, органические кислоты, аминокислоты, 0,1-2 сек, большая часть метки обнаруживалась в 3-фосфоглицериновой кислоте. Следовательно, 3-ФГК – первичный продукт фотосинтеза.
Далее, для определения первичного акцептора углекислого газа водоросли экспонировали при высокой концентрации СО2 (1%), а затем резко ее снижали (0,003%). В условиях дефицита СО2 накопится то соединение, которое является акцептором углекислого газа. Им оказалось рибулезо-1,5-бифосфат.
Дальнейшая работа в лаборатории Кальвина и в других лабораториях привела к расшифровке всех последующих реакций С3-пути фотосинтеза, который получил название цикла Кальвина. Этот цикл состоит из 4 этапов (рис.1):
1. Стадия карбоксилирования. На этой стадии происходит карбоксилирование рибулезо-1,5-бифосфата и образование двух молекул ФГК.
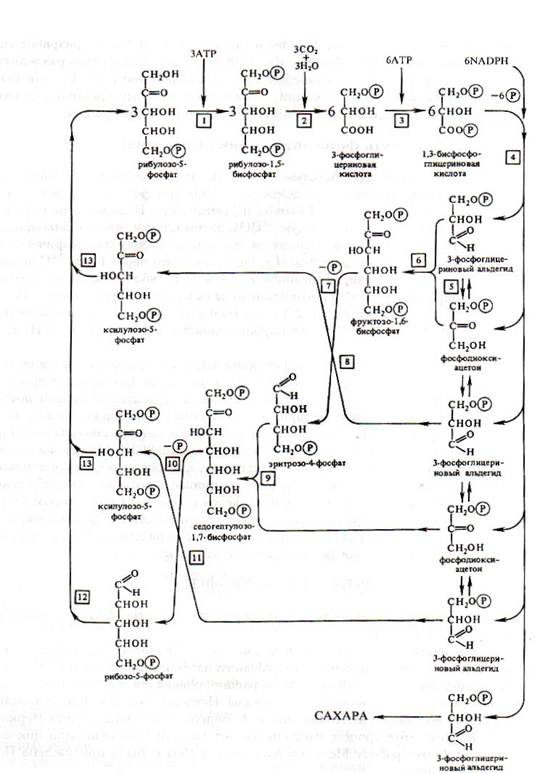
Рис 1. С3-путь фотосинтеза (цикл Кальвина).
1 – рибулозо-5-фосфаткиназа, 2 – рибулозо-1,5-бифосфаткарбоксилаза, 3 – фосфоглицераткиназа, 4 – триозофосфатдегидрогеназа, 5 – триозофосфатизомераза, 6 – альдолаза, 7 – фосфотаза, 8 – транскетолаза, 9 – альдолаза, 10 - фосфотаза, 11 – транскетолаза, 12 – рибозофосфатизомераза. 13 – фосфокетопентозоэпимераза.
2. Стадия восстановления ФГК. На этой стадии 3-ФГК восстанавливается до 3-ФГА. Этот процесс идет в два этапа: сначала 3-ФГК фосфорилируется, а затем 1,3-ФГК восстанавливаентся с участием NADPH до 3-ФГА.
3. Стадия регенерации рибулезо-1,5-бифосфата (РуБФ). Происходит с участием тре-, четырех-, пяти-, шести-, семиуглеродных сахаров. В результате процессов альдольной конденсации, катализируемых соответствующими альдолазами, и транскетолазных реакций из 5 молекул трех-углеродных сахаров образуется 5 молекул 5-углеродных сахаров, которые превращаются в РуБФ.
4. Стадия синтеза углеводных продуктов. На этом этапе из ФГА и фосфодиоксиацетона (ФДА) под действием альдолазы синтезируется фруктозо-1,6-бифосфат, который затем может превращаться или в крахмал, или в сахарозу.
Для синтеза одной молекулы глюкозы в цикле Кальвина необходимо 12 молекул NADPH, 18 молекул АТР.
2. С4-путь фотосинтеза
Листья некоторых растений (сахарного тростника, кукурузы, сорго и др.) способны фиксировать углекислый газ не только в реакциях цикла Кальвина, но и другим путем, в ходе которого появляются С4-кислоты – ЩУК, яблочная, аспарагиновая. Такой способ связывания углекислоты получил название С4-пути фотосинтеза (рис.2). Его осуществляют многие виды семейств злаковые, осоковые, амарантовые, астровые, маревые, молочайные, портулаковые и др. Данный путь фотосинтеза, отличный от С3-пути, впервые описали австралийцы М.Д.Хетч и К.Р.Слэк. Поэтому нередко этот способ связывания углекислоты называют путем Хетча и Слэка.
Для листьев С4-растений характерно анатомическое строение кранц-типа. Проводящие пучки у таких растений окружены двумя слоями зеленых клеток ассимиляционной паренхимы. Внешний слой образуют клетки-мезофилла, а внутренний слой составляют клетки-обкладки сосудистого пучка, которые также содержат хлоропласты. Клетки мезофилла и обкладки отличаются структурно и функционально. Хлоропласты клеток обкладки содержат много зерен крахмала и не имеют гран. В клетках мезофилла хлоропласты имеют обычную структуру. В клетках мезофилла происходит присоединение углекислого газа к ФЕП с образованием оксалоацетата, который далее восстанавливается до малата. Далее малат или аспартат транспортируются в клетки обкладки, где декарбоксилируются с образованием пирувата (С3-кислота). После этого С3-кислоты возвращаются в клетки обкладки (превращаются в первичный акцептор СО2), а углекислота попадает в цикл Кальвина.
С4-путь фотосинтеза характерен для растений засушливых и тропических районов, устойчивых к засухе и засолению. Растения с -путем способны осуществлять фотосинтез даже при закрытых устьицах, т.к. хлоропласты клеток обкладки используют малат (аспартат), образовавшийся ранее в клетках мезофилла, как донор СО2. Они могут также использовать углекислый газ, образовавшийся при фотодыхании. Для С4-растений характерны высокие скорости фотосинтеза (40-80 мг СО2/(дм2хч), для С3 –растений - 15-40 мг СО2 /(дм2хч). У С4-растений более высокие скорости роста - 4-5 г сухого вещества на 1 дм2 поверхности листа в день по сравнению с 0,5-2,0 г у С3-растений
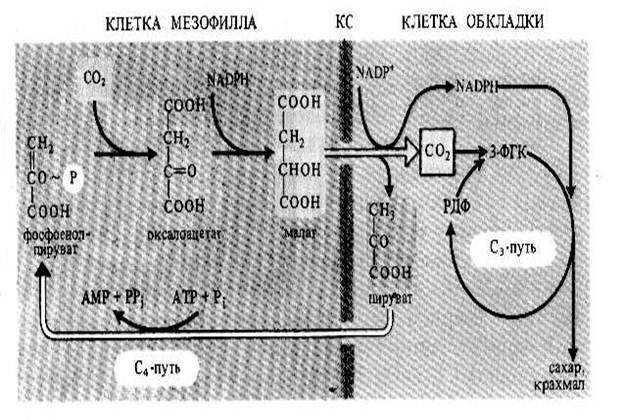
Рис.2. С4- путь фотосинтеза (путь Хэтча и Слэка)