Изменения морфологической структуры тканей
В обычных условиях посола мышечные волокна не разрушаются, в основном изменяется их толщина. Наблюдается сжатие соединительнотканных и жировых прослоек, а также капилляров кровеносной системы. Диаметр мышечных волокон на вторые сутки посола сокращается, но после четырех-пяти суток начинает возрастать, а к концу процесса он несколько превышает начальную величину. В циркулирующем рассоле мышечные волокна сжимаются, появляется грубая поперечная складчатость.
В процессе посола отчетливо изменяется внутренняя структура мышечных волокон. Уже через 12 ч поперечная исчерченность становится менее заметной, а кое-где исчезает. В дальнейшем наблюдается разрушение и слияние в аморфную массу белков миофибрилл. При посоле с вибрацией очень быстро происходит поперечный разрыв мышечных волокон на звенья, по длине близкие к диаметру волокон [10]. При посоле шкур толщина эпидермиса в связи с потерей воды несколько уменьшается. Толщина дермы резко сокращается и составляет после тузлукования в течение одних суток менее 2/3 первоначальной. Толщина сосочкового слоя также становится меньше. Наклон корней волос увеличивается. Все это указывает на значительное уплотнение слоя дермы, которое отражается и на форме потовых желез. В результате на микрофотографиях тузлукованных шкур потовые железы почти не обнаруживаются. Сальные железы, наоборот, почти не изменяются, они только уплотняются и ближе прилегают к корням волос.
Наиболее важные изменения у основного сетчатого слоя дермы. Если в срезах парной шкуры среди пучков коллагеновых волокон дермы часто заметны промежутки и пустоты, то в срезах, полученных из консервированных шкур, пучки коллагеновых волокон расположены чрезвычайно плотно. Это уплотнение дермы вызывается, по-видимому, не только обезвоживанием тканей, но и некоторым разбуханием коллагеновых волокон, так как в срезах после посола они имеют несколько большую толщину.
1. Химические изменения
Химические изменения мяса в процессе посола обусловлены по преимуществу деятельностью ферментов и лишь в небольшой части имеют спонтанный характер (например, образование азотистой кислоты из нитритов в присутствии ионов водорода). Какие ферменты, микробиальные или тканевые, играют основную роль, еще не выяснено. По-видимому, все же, ферменты, выделяемые микроорганизмами, имеют для развития некоторых процессов решающее значение. Об этом свидетельствует, во-первых, то, что в отличие от тканевых, микробиальные ферменты постоянно воспроизводятся в ходе обмена веществ микробных клеток и, следовательно, они количественно и качественно меняются в процессе посола. Во-вторых, экспериментально доказано, что присутствие микрофлоры влияет на конечное состояние продукта.
Скорость химических изменений мяса в процессе посола сравнительно невелика, так как он обычно производится при низких плюсовых температурах (порядка —4 — 0°С). Однако продолжительность посола в большинстве случаев достаточна для того, чтобы эти изменения оказались ясно выраженными. И, если ставится задача получить продукт с определенными качественными характеристиками, необходимо обеспечить достаточную продолжительность пребывания мяса в посоле.
К числу наиболее важных химических изменений мяса в процессе посола относятся: изменения составных частей мяса, изменения связанные с сохранением окраски, присущей мясу.
2. Изменение составных частей мяса
В результате непрекращающейся деятельности тканевых ферментов и ферментов, выделяемых микроорганизмами, некоторое количество белковых веществ мяса подвергается гидролитическому распаду.
При посоле, таким образом, белковые вещества теряются не толькоза счет перехода в рассол, но и вследствие распада. В табл. 84 приведены результаты опытов по посолу небольших кусков мяса в 18%-ном рассоле, из которых можно получить представление о динамике распада белков.
Таблица 84
| Содержание белкового азота к исходному, % | Продолжительность посола, сутки | |||
| В мясе | 88,1 | 83,2 | 79,1 | |
| В рассоле | 3,9 | 5,2 | 6,8 | |
| Всего | 92,0 | 88,4 | 85,9 |
По истечении 40—45 суток уменьшается даже количество общего азота, что свидетельствует о минерализации органических азотистых веществ.
В производственных условиях при посоле сравнительно крупных отрубов степень гидролитического распада белков меньше, но все же она заметна и возрастает в зависимости от продолжительности посола. По данным ВНИИМПа, при посоле окороков на 52-е сутки количество аминокислот составляло около 60, а количество аммиака около 16 мг на 100 г средней пробы мяса.
Более глубокие изменения претерпевают экстрактивные вещества мяса. Так, количество органических фосфорных соединений на 30-е сутки посола уменьшается примерно вдвое в сравнении с первоначальным. Эти изменения касаются преимущественно растворимых фосфорных соединений и почти не затрагивают нуклеопротеидов и фосфатидов. Количество креатина падает в 10—20 раз.
В результате сложных и еще мало изученных изменений, происходящих при посоле, рН мяса к концу посола обычно достигает 6,0—6,4. Если рН окажется меньшим, это указывает на закисание мяса под действием микробов, выделяющих кислоты; если же он окажется выше 6,8 — на распад белков под действием гнилостных микроорганизмов. При консервировании шкур рН рассола (тузлука) должен быть более 6,0 и менее 7,0. При рН менее 6,0 происходит кислотное набухание дермы (нажор), шкура оказывается чрезмерно влажной. При рН больше 7,0 быстрее развивается микрофлора.
Изменения белковых и других составных частей мяса, в том числе, по-видимому, и жиров, при посоле имеют и другое, положительное значение. Благодаря им соленый продукт приобретает более мягкую консистенцию и специфические приятные запах и вкус, которые зависят от вида мяса. Особенно ясно выражено изменение запаха и вкуса при посоле свинины, приобретающей свойства так называемой ветчинности.
Процесс образования ветчинности мало изучен. Установлено, что своеобразный аромат и вкус ветчинности появляется в обычных условиях посола примерно на 10—14-е сутки посола, ясно выражены к 21-м суткам, наибольшей интенсивности достигают после 50—60 суток посола. Они более ярко выражены, если посол вначале производится в рассоле и заканчивается вне рассола. Так, специфические запах и вкус ветчинности более заметны, если окорок находится 14 суток в рассоле и 7 суток на стекании, чем все 21 сутки в рассоле.
При посоле измельченной и тщательно промытой рассолом свинины аромат и вкус ветчинности не обнаруживаются. При посоле экстракта из этого мяса сравнительно быстро появляются признаки ветчинности. Стало быть, решающая роль в образовании ветчинности принадлежит экстрактивным веществам мяса.
Признаки ветчинности в значительной мере являются результатом каких-то вторичных превращений составных частей мяса, происходящих при тепловой обработке соленого продукта.
Созревшая свинина приобретает ветчинность раньше, чем несозревшая. Повышение температуры посола существенным образом ускоряет этот процесс. При посоле свинины в условиях, близких к асептическим, ветчинность появляется с большим запозданием или совсем не обнаруживается. Ветчинность обнаруживается в рассоле и мясе одновременно с появлением микроорганизмов, способных продуцировать ароматические вещества.
Таким образом, очевидно, что приобретение продуктом ветчинности в процессе посола есть результат совместной деятельности тканевых ферментов и микроорганизмов в присутствии поваренной соли. Чем меньше концентрация соли, тем яснее выражены признаки ветчинности. Однако, если концентрация соли в рассоле слишком низка (менее 10—12%), не происходит подавления гнилостных бактерий и мясо портится.
Изменение окраски мяса. Естественная окраска мяса обусловлена главным образом наличием в мышечной ткани миоглобина, составляющего около 90% общего количества пигментов мяса, и лишь отчасти наличием гемоглобина.
Простетическая группа гемоглобина и, особенно миоглобина — гем, легко окисляется, отдавая один электрон. Возможны два типа окисления гема: окисление двухвалентного железа гема до трехвалентного с образованием коричневого метмиоглобина (метгемоглобина) и окисление порфиринового кольца гема. Окисление порфиринового кольца вначале приводит к разрыву a -метинового мостика в кольце
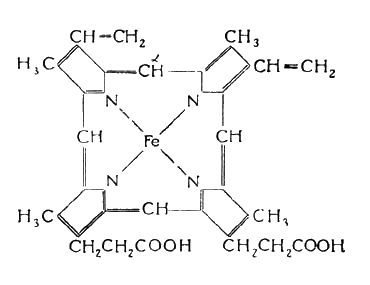
В результате образуется зеленый пигмент. При более глубоком окислении может происходить разрыв и других метиновых мостиков порфириновой системы с образованием различных продуктов окисления с разнообразной окраской. В их числе могут присутствовать зеленые производные гематина: холеглобин, вердоглобин, зеленый гемин. Если разрушение кольца сопровождается отщеплением железа, образуется желтый биливердин.
Употребляемый при посоле хлористый натрий ускоряет окисление гемовых пигментов. И хотя это окисление ограничивается в основном образованием метпигментов, естественная окраска мяса сравнительно быстро исчезает и оно приобретает коричневато-серый цвет с различными оттенками.
В большинстве случаев соленые изделия подвергают тепловой обработке. Происходящая при этом денатурация гемовых пигментов сопровождается отщеплением гема. Координационная структура этого комплекса ненасыщена. Благодаря этому гем может связывать органические основания, присоединяющиеся к центральному атому железа побочными валентностями:

Так, если атом железа в комплексе двухвалентен, образуются гемохромогены.
Двухвалентное железо гемохромогенов более чувствительно к окислению, чем двухвалентное железо гемо- и миоглобина. В связи с этим лабильно связанный кислород оксигемомиоглобина, легко отщепляющийся при его денатурации, частью окисляет железо гемохромогенов до трехвалентного, частью воздействует на порфириновое кольцо гемохромогенов. Окисление двухвалентного железа до трехвалентного приводит к переходу гемохромогенов в парагематины, обладающие различной окраской. Это усиливается в условиях контакта с кислородом воздуха.
Превращения гемовых пигментов, вызываемые их денатурацией, и частичное окисление порфиринового кольца в процессе тепловой обработки мяса приводят к полному исчезновению естественной окраски даже в том случае, если часть пигментов при посоле осталась не окисленной.
Во избежание нежелательных изменений окраски в практике посола мясо и мясопродукты обрабатывают нитритами (преимущественно натрия) и нитратами (преимущественно калия). В обоих случаях в конечном счете образуются ярко-красные нитрозопроизводные (азоксипроизводные) тех или иных исходных веществ: нитрозогемомиоглобин (азоксигемомиоглобин), нитрозогемохромогены (азоксигемохромогены), нитрозопарагематины (азоксипарагематины). Образование этих нитрозопроизводных происходит в результате взаимодействия исходных веществ с окисью азота, выделяемой нитратами и нитритами в условиях посола, по схеме
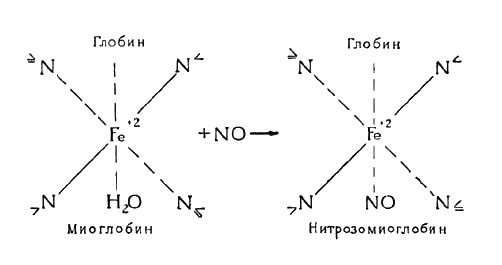
Нитрозопигменты образуются только в отсутствии кислорода. Достаточные их количества накопляются в мясе в пределах рН 5,2—6,6. При рН 7,0 и выше их количество незначительно.
Эффект стабилизации окраски мяса при посоле находится в зависимости от хода основных звеньев общей цепи химических превращений, осложняемых деятельностью денитрифицирующих бактерий: восстановления нитратов до нитритов; образования азотистой кислоты из нитритов; восстановления азотистой кислоты до окиси азота; взаимодействия окиси азота с гемовыми пигментами; сопутствующих химических процессов, ухудшающих условия стабилизации окраски.
Восстановление нитратов до нитритов под влиянием жизнедеятельности денитрифицирующих бактерий подробно описано в главе III. Реакция денитрификации может происходить в пределах рН 5,6—9,0. При оптимальном для денитрификации значении рН (7,0—8,0) образуются преимущественно продукты более глубокой денитрификации. При крайних значениях рН (5,6 и 9,0) денитрификация идет преимущественно до нитритов, необходимых для обеспечения нормальной окраски мясопродуктов. Практическое значение имеет нижний предел, т. е. рН 5,6, поскольку щелочная среда для мясопродуктов недопустима. Он, таким образом, находится в границах оптимальных для образования нитрозопигментов значений рН.
Нитрит как соль сильного основания и слабой кислоты в присутствии воды гидролизуется

В кислой среде эта реакция сдвигается в правую сторону. Снижение рН, следовательно, способствует накоплению азотистой кислоты. Но при слишком низких значениях рН происходит чрезмерно интенсивное образование летучей азотистой кислоты, которая частично переходит в окружающую среду. К тому же эта кислота в кислой среде, особенно при повышенной температуре, неустойчива и разлагается по схеме

Наряду с окисью азота образуется сильный окислитель, двуокись азота, вызывающая нежелательные окислительные изменения гемовых пигментов. Поэтому значения рН ниже5,5 не рекомендуются.
В восстановительной среде, т. е. в присутствии донаторов электронов (восстановителей), азотистая кислота восстанавливается до окиси азота
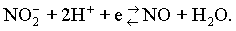
Оптимальное значение рН для этой реакции находится в границах рН 5,7—5,9 [22], т. е. в границах оптимальных значений рН для образования нитрозопигментов.
При одновременном отсутствии и восстановителей, и окислителей (например, кислорода воздуха) реакция протекает с образованием и окиси, и двуокиси азота
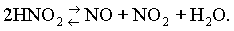
Тогда наряду с нитрозопигментами образуются и метпигменты

Именно с этим связано приобретение мясом коричневого оттенка вначале при посоле с применением нитрита, так как возникает локальный избыток его в сравнении с количеством редуцирующих веществ в мясе. Аналогичная картина и по той же причине может получиться при употреблении чрезмерно больших количеств нитрита.
Из метпигментов могут образовываться нитрозопигменты лишь после предварительного их восстановления. Здесь оказывают влияние редуцирующие вещества, повышенная температура и наличие микробиальных ферментов — нитритредуктаз (например, в период обжарки колбасных изделий). В присутствии кислорода окись азота взаимодействует с ним, превращаясь в двуокись азота со всеми вытекающими отсюда последствиями.
Таким образом, эффективность стабилизации окраски зависит от присутствия в мясе редуцирующих веществ, а повысить скорость окрашивания, интенсивность окраски и ее устойчивость можно, если ввести одновременно с нитритом восстановители.
Однако все эти процессы осложняются многочисленными побочными явлениями, большинство которых еще недостаточно выяснено. Во всяком случае вследствие высокой реакционной способности азотистой кислоты не исключаются процессы ее взаимодействия с рядом азотистых веществ, входящих в состав мяса [42], в том числе с аминокислотами и высокомолекулярными азотистыми веществами (полипептидами, белками). При этом часть нитрита расходуется на эти реакции, а вещества, с которыми они взаимодействуют, изменяясь, утрачивают свои свойства. Правда, в количественном отношении эти потери не имеют существенного значения.
В отличие от свежего мяса пигменты соленого мяса нитрозомиоглобин и нитрозогемоглобин более чувствительны к окислению, в особенности на свету [25]. Поверхность соленого продукта изменяет окраску в темноте через несколько суток, на свету — через несколько часов. Окисление сопровождается образованием метпигментов и разрушением порфиринового кольца. Поверхность мяса светлеет, приобретает серый, коричневатый оттенок, местами зеленую или желтоватую окраску.
Часть этих изменений связана с воздействием на продукт кислорода воздуха, под влиянием которого нитрозопигменты переходят в метпигменты

Образующаяся при этом окись азота в свою очередь вызывает более глубокие окислительные изменения простетической группы.
При постоянном освещении происходит окисление кислородом свободных сульфгидрильных групп белков (в особенности, денатурированных). При этом как промежуточный продукт образуется перекись водорода, которая, взаимодействуя с окисью азота, способна давать пероксид азотистой кислоты. Последний вызывает окислительную деструкцию гемовых пигментов до зеленых биливердиновых пигментов [42]. Аналогичный процесс возникает и в случае возникновения перекиси водорода в результате жизнедеятельности некоторых бактерий, которые хотя и погибают при нагреве до 70° С, но ферменты их сохраняют активность (бактериальное позеленение) [32]. Окись азота и пероксид азотистой кислоты при сухом посоле способствуют также и окислению жиров [42].
Вероятность возникновения пороков окраски готового продукта уменьшается применением при посоле сильных восстановителей: аскорбиновой, никотиновой, сернистой кислот, цистеина, глютатиона [32] и, отчасти, редуцирующих сахаров (глюкозы, инвертного сахара). Вместе с тем эти вещества ускоряют процесс цветообразования.
Чаще всего применяют аскорбиновую кислоту, которая реагирует непосредственно с азотистой кислотой, образуя окись азота и дегидроаскорбиновую кислоту
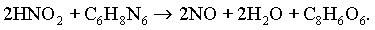
Эта реакция сравнительно медленно протекает при низких температурах, но резко ускоряется при обжарке и копчении.
Аскорбиновая кислота, кроме этого, способна восстанавливать метпигменты, в том числе и денатурированные
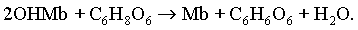
Она снижает парциальное давление кислорода в окружающем воздухе и окислительно-восстановительный потенциал в соленом продукте, стабилизируя этим его окраску [42], но практически не восстанавливает нитратов.
В практике пользуются и аскорбиновой кислотой и ее солями — аскорбинатами. Последние применять при посоле удобнее, так как при их совместном растворении с нитритом не возникает немедленного взаимодействия с ним, поскольку среда слабощелочная, в то время как аскорбиновая кислота немедленно вступает в реакцию с нитратом.
Однако при чрезмерно больших количествах аскорбиновой кислоты может произойти ухудшение окраски. Часть аскорбиновой кислоты при высокой концентрации, окисляясь, отдает электроны пигментам. Образуется неустойчивая гидроперекись гемовых пигментов, распадающаяся на метпигмент и холеглобин.
Дозировка аскорбиновой кислоты 47 г, а аскорбината натрия 52 г на 100 кг мяса (с некоторым избытком). Избыток разрушается в период тепловой обработки, так что в готовом продукте остается 0—7 г аскорбината на 100 кг продукта. При посоле мяса аскорбинат добавляют к шприцовочному рассолу, исходя из приведенной выше нормы. Фосфаты усиливают эффект действия аскорбиновой кислоты. Предполагается, что они играют роль синергистов восстановителей [22, 38]. В наибольшей степени эти свойства выражены у триполифосфата.
Вопрос о роли сахара в стабилизации окраски соленых продуктов нитратом и нитритом еще не вполне ясен. Часть исследователей не придают ему практического значения [24]. Другие придерживаются обратного мнения [39]. Судя по всему, однако, добавление сахара в посолочные смеси не может не влиятьвтой или иной форме на цветообразование.
При посоле различных изделий из мяса часто наряду с солью и нитратом, нитритом или их смесью сахар употребляют преимущественно в виде сахарозы. Основная цель — смягчение вкуса солености, для чего необходимо 1,5—2,5% сахара к весу сырья (в зависимости от солености). Сахар более быстро и равномерно распределяется в тканях, чем соль. К концу посола в рассоле остается 30—40% сахара к его начальному количеству. Часть его (25—55%) переходит из рассола в мясо. Остальное количество исчезает. Это обусловлено жизнедеятельностью микроорганизмов, о чем говорилось в главе III.
В процессе посола некоторая, иногда значительная часть сахара (до 60%) в ходе разложения подвергается инверсии под влиянием воды, кислот и инвертазы бактерий, образуя моносахариды — глюкозу и фруктозу. Степень инверсии зависит от продолжительности посола. Таким образом, на какой-то стадии посола, если он достаточно продолжителен, даже в случае применения дисахаридов, в мясе и рассоле накапливаются моносахариды. Если же при посоле употребляют не сахарозу, а моносахариды (глюкозу, инвертный сахар), количество моносахаридов с самого начала будет значительным.
Как было показано в главе III, моносахариды, т. е. редуцирующие сахара, являются донаторами электронов и, следовательно, участниками процесса денитрификации. Вследствие этого добавление к посолочной смеси моносахаридов несколько ускоряет денитрификацию. Добавление сахарозы сказывается на скорости денитрификации очень мало, поскольку в данном случае необходима предварительная инверсия сахара.
Дыхание некоторых денитрифицирующих микроорганизмов за счет использования моносахаридов идет интенсивнее, чем за счет восстановления нитритов до нежелательной степени. Поэтому в присутствии моносахаридов процесс разрушения азотистой кислоты тормозится [24]. Это делает окраску более интенсивной и устойчивой. Если поверхность разреза колбасы обработать раствором моносахарида, она дольше сохраняет окраску на свету. Свинина, посоленная с глюкозой, дольше сохраняет окраску, чем без нее.
Наконец, нельзя исключить вероятности прямого взаимодействия азотистой кислоты как акцептора и моносахаридов как донаторов электронов по схеме
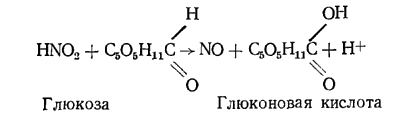
Иначе говоря, в присутствии моносахарида уменьшается вероятность разложения азотистой кислоты с образованием двуокиси азота.
С способностью моносахаридов служить донаторами электронов связана, по-видимому, и их способность частично восстанавливать метпигменты [39].
Все это дает основание оценивать положительно роль моносахаридов в ускорении, улучшении и повышении стабильности окраски соленых мясопродуктов. Количества моносахаридов, достаточные для этих целей, составляют 0,2—0,3% к весу мяса [39]. Что касается сахарозы, то ее влияние в незначительной степени проявляется лишь при длительном посоле.
В тесной связи с содержанием в рассоле и мясе редуцирующих веществ находится допустимое количество нитрита. При его большом избытке относительно количества редуцирующих веществ идет разложение азотистой кислоты. Это приводит к переходу гемовых пигментов в метпигменты и может сопровождаться окислением порфиринового кольца. Красная окраска соленого продукта местами исчезает, сменяясь серой, буроватой, а иногда зеленоватой. Свинина, посоленая в рассоле, содержащем более 0,1% нитрита, частью может приобретать зеленоватый оттенок. То же произойдет и с самим рассолом при его длительном употреблении.
Для предотвращения обесцвечивания мяса можно употреблять как нитрит, так и нитрат. Пользование нитритом удобнее в том отношении, что должный эффект достигается значительно быстрее. Но окраска, достигнутая с помощью нитрита, неустойчива. Неустойчивость окраски обусловлена, во-первых, окислительным воздействием кислорода воздуха, который способен проникать на глубину до 2 мм, во-вторых, деятельностью микробов. Некоторые из них продуцируют перекись водорода, способную окислять окись азота. Другие образуют сероводород, который в присутствии кислорода дает с миоглобином зеленый сульфмиоглобин. Повышение устойчивости окраски за счет больших количеств нитрита недопустимо не только из-за ухудшения окраски, но и вследствие опасности отравления, так как нитрит ядовит. Его единовременная фармакологическая доза составляет около — 0,3 г, суточная — 1,0 г. В связи с этим санитарным законодательством СССР установлена предельно допустимая норма содержания нитрита в пищевых продуктах, а именно 20 мг нитрита натрия на 100 г продукта.
Вследствие неустойчивости окраски, получаемой с помощью нитрита, его применяют лишь в тех случаях, когда продолжительность пребывания продукта в производстве невелика, а готовый продукт не предназначен для длительного хранения (например, вареные колбасные изделия). Минимальная доза нитрита натрия, достаточная для получения удовлетворительного результата, около 0,005% (5 мг на 100 г мяса) [6]. Она была установлена экспериментально, но, по-видимому, близка к теоретически необходимому количеству.
Дозировка нитрита при посоле рассолами зависит от вида мяса. ВНИИМПом рекомендуется при посоле говядины и баранины 0,10% и при посоле свинины 0,06% к весу рассола.
Для сохранения окраски изделий, находящихся в производстве длительное время или предназначенных для длительного хранения (соленые штучные изделия, сырокопченые колбасы и пр.), пользуются нитратами. Так как нитраты не столь токсичны, как нитриты, их доза может быть большей: 0,05—0,10% к весу мяса, т. е. во много раз больше, чем необходимо для образования нужного количества нитрита. Благодаря этому нитрат длительное время служит источником пополнения той части нитрита, которая разлагается под влиянием тех или иных причин.
Количество нитрита, которое получается из нитрата в процессе посола, зависит от условий, в которых он протекает, и длительности его. Скорость образования нитрита в течение 30—40 суток невелика, а затем резко возрастает. Однако при нормальных условиях посола его количество в продукте по окончании посола не превышает установленной законом нормы и составляет не более 14 мг при мокром и не более 16 мг на 100 г продукта при сухом посоле. В литературе, однако, встречаются указания на то, что употребление чрезмерно больших количеств нитрата может привести к накоплению в мясе опасных количеств нитрита [35].
В связи с тем, что скорость образования нитрита из нитрата меньше скорости диффузии соли в тканях, время, необходимое для приобретения продуктом желательной окраски, оказывается более продолжительным, чем для его просаливания. Отсюда длительность посола с применением нитрата иногда определяется не скоростью собственно посола, а скоростью окрашивания мяса. Поэтому при сокращенных сроках посола для предохранения мяса от обесцвечивания следует употреблять смесь нитрата с нитритом. Дозировка нитрата и нитрита в этом случае зависит от способа и условий посола.
Значение нитратов и нитритов при посоле не исчерпывается их влиянием на окраску мяса. В отсутствии нитратов и нитритов характерный аромат ветчинности мало заметен. Это заставляет связывать появление аромата ветчинности с образованием каких то продуктов превращения нитратов и нитритов в связи с развитием жизнедеятельности некоторых бактерий.
Вместе с этим нельзя отрицать и участия в сложных процессах образования признаков ветчинности экстрактивных веществ мяса. Аромат и вкус соленых изделий зависят от вида мяса: соленые изделия из свинины по аромату и вкусу значительно отличаются от соленых изделий из говядины и баранины.
Длительные наблюдения показали, что при посоле с применением нитратов развитие признаков ветчинности и развитие окраски близко совпадают по времени.