ЛЕКЦИЯ 3. ФИЗИОЛОГИЯ МЫШЦ
План
1. Морфофункциональная организация мышечного волокна.
2. Нервно-мышечная передача.
3. Механизм мышечного сокращения и расслабления.
Морфофункциональная организация мышечного волокна. Структурной и функциональной единицей скелетной мышцы является мышечное волокно. Оно представляет собой вытянутую клетку диаметром около 10-100 мкм и длинной 10-12 см. Мышечное волокно окружено клеточной мембраной – сарколеммой. В его саркоплазме содержатся ядра, клеточные органоиды и сократительные элементы - миофибриллы. Миофибриллы представляют собой длинные нити диаметром 1-2 мкм, сгруппированные в пучки по 4-20 штук в каждом. Они состоят из тонких и толстых протофибрилл (миофиламентов). Тонкие протофибриллы состоят из белка актина, а толстые – из белка миозина. Актиновые нити прикреплены к особым белковым образованиям -  - пластинкам. Толстые миозиновые нити лежат между актиновыми. Участок между двумя - пластинками называют саркомером.
- пластинкам. Толстые миозиновые нити лежат между актиновыми. Участок между двумя - пластинками называют саркомером.
Актин составляет 20 % сухого веса миофибрилл. В мышечном волокне актин находится в фибриллярной форме (F-актин). Волокна фибриллярного актина выглядят как две нити, закрученные одна вокруг другой. Миозин составляет 50-55 % от сухой массы миофибрилл и состоит из уплощенных параллельно белковых нитей. На обоих конках его имеются отходящие в стороны шейки с уплощениями - головками. Благодаря головкам образуются поперечные мостики между актином и миозином. Вдоль актиновых нитей миофибрилл расположены регуляторные субъединицы, состоящие из белкой тропонина и тропомиозина. Эти белки в невозбужденной мышце тормозят взаимодействие между актином и миозином.
Специфическими элементами мышечного волокна является система продольных трубочек – саркоплазматический ретикулум и система поперечных трубочек – Т-система, представляющая собой впячивания поверхностной мембраны. В каналах саркоплазматического ретикулума содержится избыточное количество ионов Са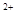 .
.
Нервно-мышечная передача.Основная функция скелетной мышцы – сокращение. Для реализации этой функции необходимо, чтобы возбуждение перешло с нерва на мышцу. Это происходит через нервно-мышечный синапс. Нервно-мышечный синапс состоит из мембраны нервного окончания (пресинаптической мембраны), мембраны мышечного волокна (постсинаптической мембраны) и расположенной между ними синаптической щели. В нервно-мышечных синапсах возбуждение передается химическим путем. Медиатором является ацетилхолин. Он находится в пузырьках расположенных в окончании двигательного нерва. Постсинаптическая мембрана содержит холинорецепторы и фермент холинэстеразу, способный разрушать ацетилхолин.
Механизм передачи возбуждения состоит в следующем. Нервные импульсы проходящие по двигательным волокнам, производят деполяризацию мембраны нервного окончания, что вызывает разрушение оболочки пузырьков и поступление в синаптическую щель ацетилхолина. Молекулы ацетилхолина диффундируют к постсинаптической мембране мышечного волокна и связываются с холинорецепторами мембраны. Это приводит к повышению проницаемости постсинаптической мембраны для Na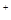 и К. Положительно заряженные ионы устремляются внутрь мышечного волокна и на их мембране возникает постсинаптический потенциал или потенциал концевой пластинки. Создавшаяся разность потенциалов между постсинаптической мембраной и мембраной мышечного волокна создает местный ток, возбуждающий мышечную мембрану. В ней возникает потенциал действия, распространяющийся по мышечному волокну. Выделившийся ацетилхолин разрушается ферментом холинэстеразой, и постсинаптическая мембрана приобретает свой исходный заряд.
и К. Положительно заряженные ионы устремляются внутрь мышечного волокна и на их мембране возникает постсинаптический потенциал или потенциал концевой пластинки. Создавшаяся разность потенциалов между постсинаптической мембраной и мембраной мышечного волокна создает местный ток, возбуждающий мышечную мембрану. В ней возникает потенциал действия, распространяющийся по мышечному волокну. Выделившийся ацетилхолин разрушается ферментом холинэстеразой, и постсинаптическая мембрана приобретает свой исходный заряд.
Механизм мышечного сокращения и расслабления.Нервный импульс распространяется вдоль мышечного волокна и заходит в поперечные трубочки внутрь волокна. Он вызывает выход из цистерн саркоплазматического ретикулума ионов Са, которые проникают к миофибриллам. Под влиянием Са длинные молекулы тропомиозона проворачиваются вдоль оси и скручиваются в желобки между молекулами актина, открывая участки прикрепления головок миозина к актину. Между актином и миозином образуются поперечные мостики . При этом головки миозина совершают гребковидные движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов к центру. При однократном движении поперечных мостиков вдоль актиновых нитей саркомер укорачивается на 1 % его длины. Для дальнейшего скольжения сократительных белков мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания ионов кальция. В этот момент происходит активация молекул миозина. Миозин приобретает свойсва фермента АТФ-азы и вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии ионов кальция новых мостиков на следующем участке актиновой нити. В результате многократного повторения подобных процессов сокращается длина саркомеров и всего мышечного волокна. Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью.
Расслабление мышцы начинается с быстрого поглощения ионов кальция саркоплазматической сетью каналов. При этом восстанавливают свои ингибирующие свойства тропонин и тропомиозин. Актомиозиновый комплекс разрушается: актин и миозин взаимно отталкиваются и мышца возвращается в исходное, расслабленное состояние.