Механизм сокращения и расслабления мышечного волокна
Управление мышцей, состоящей из значительного числа двигательных единиц (ДЕ), осуществляется совокупностью мотонейронов, иннервирующих мышцу, получившей название мотонеронный пул (МП). Известно, что мотонейроны, посылающие свои аксоны к той или иной мышце, могут располагаться не только в пределах одного сегмента спинного мозга, но и занимать соседние. Таким образом, структурно МП может быть разнесен на достаточно большое расстояние в пределах нескольких сегментов передних рогов спинного мозга. Функционально МП является конечной инстанцией, где формируется структура командных сигналов на мышцу, обеспечивающих ее включение в двигательный акт. Именно на МП осуществляется интеграция входных воздействий на мотонейроны от надсегментарных структур и от рецепторов двигательной периферии.
Основная функция МП - дозирование силы сокращения мышцы - обеспечивается двояко - частотой импульсации входящих в него мотонейорнов и количеством активированных мотонейронов данного пула.
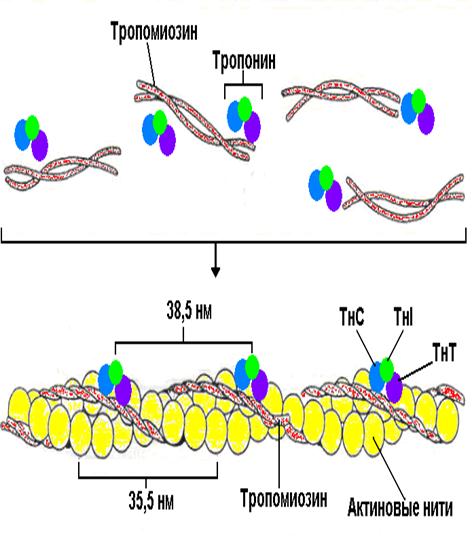
Рис. 34. Структура тонкого филамента.
Через 50 мс после произвольной внутренней команды начинается сокращение мышцы скелетной. За это время команда передается от коры к мотонейронам спинного мозга и по двигательным волокнам к мышце. Медиатором в нервно-мышечном синапсе является ацетилхолин, который содержится в синаптических пузырьках пресинапса. Нервный импульс вызывает опорожнение синаптических пузырьков и выход ацетилхолина в синаптическую щель. Там медиатор действует на постсинаптический рецептор, после чего разрушается. По мере расходования запасы ацетилхолина пополняются путем синтеза в пресинаптической мембране, но если импульсы идут часто и долго, то расход превышает пополнение и нарушается проведение возбуждения через нервно-мышечный синапс. В результате наступает утомление.
Сокращение мышцы является результатом сокращения составляющих ее мышечных клеток (волокон). Сокращение мышечного волокна – это результат укорочения каждого его саркомера за счет взаимодействия тонких и толстых филаментов.
В покоящейся мышце тонкие филаменты контактируют с Z –линиями и не достигают центра саркомера, а толстые находятся в центре, но не достигают Z-линии. Только по бокам А-диска в пространство между толстыми филаментами немного входят тонкие (Рис. 33, В).
При умеренном усилии происходит умеренное сокращение за счет того, что тонкие филаменты движутся навстречу друг другу, поэтому расстояние между Z – линиями уменьшается и длина мышцы уменьшается.
При максимальном сокращении толстые филаменты касаются Z –линий.
Физиолого-биохимический механизм сокращения состоит в том, что после взаимодействия медиатора с рецептором при достаточной частоте нервных импульсов на мышечной мембране развивается мышечный потенциал действия, который быстро распространяется вдоль мышечного волокна, вызывая выход Са2+ из саркоплазматического ретикулума. Затем Са2+ проникает в миофибриллы к центрам связывания на молекуле тропонина. В покоящейся мышце тропомиозин препятствует присоединению миозиновой головки к ближайшему с ним мономеру актина. Связывание Са с тропонином изменяет его пространственную структуру, что ослабевает связь между его тропомиозинсвязывающей субъединицей и актином. В результате молекула тропомиозина начинает двигаться по желобку тонкого филамента, освобождая скрытый до этого миозинсвязывающий центр на поверхности молекулы актина. Начинается взаимодействие актина с миозином, что сближает молекулы, принадлежащие тонким и толстым филаментам. В результате расстояние между Z-линиями уменьшается. При связывании актина и миозина молекула АТФ распадается на АДФ и фосфор неорганический. Распад АТФ ведет к расслаблению мышцы из-за изменения конформации миозина. Для восстановления способности к сокращению к головке миозина должна присоединиться следующая молекула АТФ. Весь процесс от появления мышечного потенциала до сокращения мышечного волокна называется электромеханической связью (сопряжением). Сокращение гладких мышц имеет особенность: Са2+ связывается с особым белком – кальмодулином.