Окислительное фосфорилирование. Механизм сопряжения процесса транспорта электронов с образованием АТФ. Энергетическая эффективность процесса.
Цикл Кребса.
Пути окисления органических веществ. Основные пути диссимиляции углеводов.
«Физиология растений». Дежавю. стр. 130
Цикл ди- и трикарбоновых кислот (цикл Кребса)
Аэробная фаза дыхания локализована в митохондриях. Пировиноградная кислота окисляется до воды и углекислого газа в дыхательном цикле, получившем название цикла ди- и трикарбоновых кислот или цикла Кребса в честь английского биохимика Г. Кребса, описавшего этот путь (рис 6.2.). В этом цикле окисляется не сама пировиноградная кислота, а ее производное – ацетилкоэнзим А. Он образуется в результате окислительного декарбоксилирования пировиноградной кислоты. Процесс этот состоит из ряда реакций и катализируется сложной мультиферментной системой, состоящей из трех ферментов и пяти коферментов, и названной пируваткарбоксилазой.

Рис. 6.2. Цикл Кребса (цикл ди- и трикарбоновых кислот).
1 – мультиэнзимный комплекс окислительного декарбоксилирования пировиноградной кислоты, 2 – цитратсинтаза, 3 – аконитатгидратаза, 4 – изоцитратдегидрогеназа, 5 – мультиэнзимный комплекс окислительного декарбоксилирования α-кетоглутаровой кислоты, 6 – сукцинатдегидрогеназа, 7 – фумаратгидратаза, 8 – малатдегидрогеназа (по В. В. Полевому).
При окислении одной молекулы пировиноградной кислоты образуется 3 молекулы НАДН, 1 молекула НАДФН и 1 молекула ФАДН2, при окислении которых в дыхательной электронтранспортной цепи синтезируется 14 молекул АТФ. Кроме того, 1 молекула АТФ образуется в результате субстратного фосфорилирования.
Дыхательная электронтранспортная цепь
и окислительное фосфорилирование
Дыхательная электронтранспортная цепь состоит из переносчиков электронов, которые передают электроны от субстратов на кислород. Расположение переносчиков определяется величиной их окислительно-восстановительного потенциала. Цепь начинается с НАДН, имеющего потенциал –0,32 В, и кончается кислородом с потенциалом +0,82 В. Переносчики расположены по обеим сторонам внутренней мембраны митохондрий и пересекают ее. На внутренней стороне мембраны, расположенной к матриксу митохондрии, два протона и два электрона от НАДН переходят на флавинмононуклеотид и железосерные белки. Флавинмононуклеотид, получив протоны, восстанавливается и переносит их на внешнюю сторону мембраны, где отдает протоны в межмембранное пространство. Железосерные белки, находящиеся внутри мембраны, передают электроны от НАДН окисленному убихинону Q. Он, присоединив еще два протона, диффундирует в мембране к цитохромам. Цитохром b560 отдает два электрона убихинону, который, присоединив еще два протона из матрикса, передает два электрона цитохрому b556 и два электрона цитохрому c1, а протоны выходят в межмембранное пространство. На наружной стороне мембраны цитохром с, получив два электрона от цитохрома c1, передает их цитохрому а, который переносит их через мембрану на цитохром а3. Цитохром а3, связывая кислород, отдает ему электроны. Кислород присоединяет два протона с образованием воды (рис. 6.5).
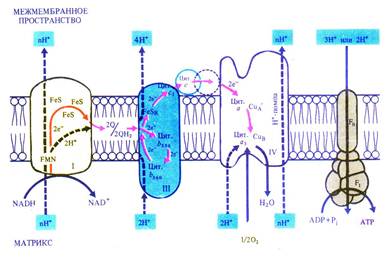
Рис. 6.5. Локализация электрон- и протонтранспортных реакций во внутренней мембране митохондрий (по В. В. Полевому).
Таким образом, транспорт электронов в дыхательной электронтранспортной цепи сопровождается трансмембранным переносом протонов. Возникающая разность потенциалов по обеим сторонам внутренней мембране митохондрий используется для синтеза АТФ (окислительное фосфорилирование), как это было показано в разделе 5.2.2. В результате прохождения двух электронов по цепи образуется 3 молекулы АТФ.