СПЕЦИАЛИЗАЦИЯ ПАТОГЕНОВ
Специализация патогена— это способность паразитировать на одном, нескольких или многих видах растений-хозяев. Специализация патогенов определяется прежде всего их приуроченностью к определенному питающему субстрату и способностью эффективно усваивать его.
В целом, картина взаимоотношений патогенов и растений-хозяев сложна и многообразна. Одни патогены способны поражать лишь определенную ткань одного вида растения-хозяина. Другие, напротив, вызывают системное поражение у видов растений, относящихся к разным классам. Между этими крайними возможностями располагается очень большое число патогенов, в разной степени поражающих разное количество видов растений-хозяев.
Различные способности патогенов к паразитизму определяются их генетическими и физиологическими свойствами. Большое значение имеют наличие генов вирулентности и эффективность их продуктов. Очень важную роль играет активность ферментативного аппарата патогенов, позволяющая использовать субстрат в доступной форме.
Специализацию патогенов определяют также защитные реакции растений-хозяев. Барьерные факторы и химические процессы блокируют внедрение многих паразитов в растение. В то же время другие паразиты способны преодолевать такие барьеры.
Нередко на специализацию паразитов оказывают влияние и внешние условия, экологические факторы. Так, в одних условиях паразит способен активно внедряться в растение и проходить в нем весь свой инфекционный цикл, в других — внедрение и развитие паразита ограничено, отдельные стадии цикла развития паразита могут выпадать. Наконец, при определенных условиях внедрение паразита в растение-хозяина не происходит. На изменение специализации патогенов может влиять хозяйственная деятельность человека.
Таким образом, специализация патогенов определяется их генетической предрасположенностью и физиологическими свойствами, эффективностью защитных реакций растений-хозяев и внешними условиями.
Понятие специализации наиболее уместно по отношению к видам патогенов.
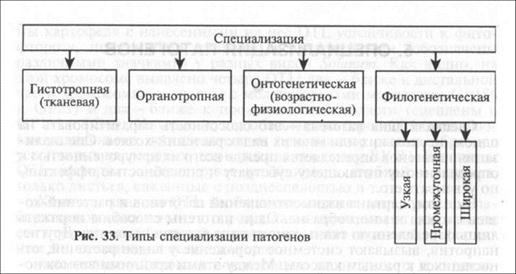
Различают несколько форм специализации патогенов к растениям-хозяевам (рис. 33).
• Гистотропная (тканевая) специализация (гис-тос — ткань) — способность патогена заселять лишь определен-
ную ткань или комплекс тканей растения. Другие ткани не поражаются. Например, возбудитель слизистого бактериоза капусты бактерия Erwinia carotovora subsp. carotovora поражает в основном паренхиму (цв. вклейка, рис. 1). Возбудитель вертицил-лезного увядания хлопчатника Verticillium dahliae поражает сосудистую систему растения (цв. вклейка, рис. 2). Риккетсии в организме растений-хозяев концентрируются преимущественно во флоэме.
• Органотропная специализация — способность патогена заселять лишь определенные органы растения. Например, Plasmodiophora brassicae поражает корни растений из семейства крестоцветных (цв. вклейка, рис. 3). Возбудитель спорыньи злаков Claviceps purpurea поражает завязи и позднее развивающиеся зерновки злаков (цв. вклейка, рис. 4).
• Онтогенетическая, или возрастно-физиологи-ческая (стадийно-возрастная) специализация-способность патогена поражать растение-хозяина в определенный период его развития. Так, возбудители корнееда сахарной свеклы Aphanomyces cochlioides и Pythium debarianum поражают проростки от момента прорастания до образования четырех настоящих листьев (цв. вклейка, рис. 5). Возбудитель твердой головни пшеницы Tilletia tritici поражает хозяина только с момента прорастания зерновки до появления всходов (цв. вклейка, рис. 6).
Многие возбудители пятнистостей заражают растения в фазе вегетации, особенно во второй ее половине. Это, например, возбудитель церкоспороза свеклы (Cercospora beticola) (цв. вклейка, рис. 7).
Возбудители пыльной головни пшеницы (Ustilago tritici) и спорыньи ржи (Claviceps purpurea) поражают растения-хозяева в фазе цветения (цв. вклейка,.рис. 8).
Наконец, некоторые возбудители способны поражать растение-хозяина на разных стадиях развития. Классический пример — возбудитель пузырчатой головни кукурузы (Ustilago zeae), способный поражать кукурузу практически в любой фазе развития растения. Дейтеромицет Botrytis cinerea поражает свои многочисленные растения-хозяева в фазах вегетации, цветения и плодоношения (последние две фазы — предпочтительнее).
• Филогенетическая специализация — способность вида патогена поражать один, несколько или много видов растений-хозяев. Филогенетической ее называют потому, что способность поражать растения-хозяева — основное направление микроэволюционного процесса паразитов. На повышение эффективности паразитизма в популяциях патогенов работает отбор, в результате действия которого появляются более приспособленные, более вирулентные формы паразитов. Результатом служит либо повышение эффективности паразитизма на одном растении-хозяине, либо расширение круга видов растений-хозяев.
Филогенетическая специализация может быть узкой и широкой.
• Узкая специализация. Патогены с узкой специализацией (их называют монофаги) паразитируют на растениях, относящихся к одному виду или роду. Узкоспециализированные патогены — это большинство облигатных паразитов и факультативных сапротрофов, а также некоторые факультативные паразиты. Они хорошо и избирательно приспособлены для поражения конкретных (нередко близкородственных) видов растений-хозяев. При коэволюции у них вырабатываются механизмы атаки, учитывающие защитные реакции, присущие видам именно их растений-хозяев.
Виды паразитов с узкой специализацией имеют выраженную иерархическую структуру (табл. 5). Обычно вид способен поражать несколько родов или видов растений-хозяев, нередко близкородственных. Вид паразита подразделяется на несколько или много разновидностей или специализированных форм (formae speciales). Они приурочены к определенному роду или виду растения-хозяина. В пределах разновидности патогена выделены физиологические расы. Они отличаются друг от друга способностью по-разному поражать сорта вида растения-хозяина. При взаимодействии определенных рас и сортов в одних комбинациях наблюдают реакцию сверхчувствительности (гибель патогена, образование мелких некрозов в месте внедрения), в других — реакцию совместимости (развитие патогена, интенсивное образование пустул).

* Разновидности выделяют, например, у Phytophthora megasperma и цветкового паразита Viscum album (омелы белой).
Ржавчинный гриб Puccinia graminis поражает злаки — хлебные, кормовые и дикорастущие. Этот вид имеет комплексную структуру. Установлено, что Puccinia graminis состоит из ряда специализированных форм, поражающих определенный род растений-хозяев. P. graminis f. sp. tritici поражает пшеницу, P. graminis f. sp. avenae — овес, P. graminis f. sp. secalis — рожь, P. graminis f. sp. poae — мятлик, P. graminis f. sp. agrostis — полевицу. Расы P. graminis отличаются друг от друга по растениям-хозяевам, молекулярным маркерам и иногда — по строению урединиоспор.
Строго говоря, специализация Puccinia graminis и других ржавчинных грибов с полным циклом распространяется даже на отдельные спороношения гриба. Так, именно урединиоспоры и эци-оспоры P. graminis заражают роды злаков, а базидиоспоры — представителей семейства Барбарисовых барбарис и магонию.
Гриб Erysiphe graminis вызывает мучнистую росу у растений из семейства Злаковых. Как и у возбудителя стеблевой ржавчины, у Erysiphe graminis имеется ряд специализированных форм, приуроченных к определенным родам злаков. Сходным образом Erysiphe labiatarum, включающий в себя много специализированных форм, паразитирует на представителях семейства Губоцветных.
Цветковое растение-паразит Viscum album (омела белая) в Европе представлено тремя разновидностями — V. album var. mali (в основном на яблоне, реже на других лиственных породах), V. album var. abietis (на пихте), V. album var. pini (на сосне). У цветкового растения-паразита Orobanche ситапа (заразиха подсолнечниковая) обнаружили три расы: А, Б и В. К расе А были созданы устойчивые сорта, ответом паразита стало появление расы Б, которая в массовом количестве поразила многие сорта подсолнечника, успешно внедряясь в его корневую систему. Позднее была выявлена и раса В, также характеризующаяся высокой вредоносностью.
Многие виды патогенов поражают только один род растения-хозяина. Возбудитель твердой головни Tilletia tritici поражает только пшеницу, а возбудитель пузырчатой головни Ustilago zeae — кукурузу. Ржавчинный гриб Puccinia menthae поражает только мяту. Цветковое растение-паразит заразиха Orobanche owerinii паразитирует только на шалфее лесном (Salvia silvestris).
• Широкая специализация. Патогены с широкой специализацией (их называют полифаги) паразитируют на растениях одного и нескольких семейств или даже различных классов. Большинство широкоспециализированных патогенов — факультативные паразиты, однако среди них встречаются и облигатные (например, некоторые ржавчинные грибы). Значительная часть факультативных паразитов соответствует формальным родам несовершенных грибов.
Plasmodiophora brassicae вызывает килу у растений из семейства Крестоцветных. Наиболее часто поражается капуста, однако могут поражаться редька, репа и другие виды этого семейства. Ржавчинный гриб Puccinia malvacearum поражает роды растений из семейства Мальвовых, оомицет Peronospora parasitica — роды растений из семейства Крестоцветных. Аскомицет Claviceps purpurea вызывает спорынью ржи и многих луговых и газонных трав из семейства Злаковых.
Botrytis cinerea поражает растения из семейств Розовых, Виноградовых, Лилейных, Амарилисовых, Пасленовых, Бобовых и др., вызывая у них серую гниль (цв. вклейка, рис. 9). Sclerotinia sclerotiorum поражает растения из семейств Зонтичных, Лилейных, Злаковых, Сложноцветных, вызывая у них белую гниль. Следует отметить, что у различных растений-хозяев при поражении этими гнилями проявляются довольно сходные симптомы поражения — мощный некроз и налет, иногда сопровождающиеся образованием склероциев разных размеров.
Phytophthora cactorum поражает растения из семейств Кактусовых, Розовых, Бобовых, Лилейных, Сосновых и др. (всего — не более 40 семейств). Особой приуроченности к органу или возрастной фазе растения гриб не имеет. Так, он вызывает корневые гнили всходов сосен и бобовых, гнили основания стебля лилий, гнили коры и плодов у розовых, гнили сердцевины и почек у рододендронов и пионов.
Ph. cinnamomi поражает в основном тропические растения — авокадо, корицу, орхидеи. Однако хозяйственная деятельность человека привела к расширению специализации этого паразита. В США и Новой Зеландии Ph. cinnamomi вызвала корневую гниль сеянцев сосны, в Европе — чернильную болезнь (поражение камбия) каштана, в Австралии — гибель эвкалиптовых лесов на больших площадях. Этот гриб поражает растения из 50 семейств Папоротникообразных, Голосеменных и Цветковых растений.
Бактерия Pseudomonas solanacearum вызывает сосудистый бактериоз растений из семейств Пасленовых, Тыквенных, Бобовых и Сложноцветных.
Цветковое растение Orobanche aegyptiaca (заразиха египетская) паразитирует на растениях из семейств Пасленовых, Тыквенных, Сложноцветных. Эта патосистема сложилась давно и остается устойчивой.
• Промежуточные случаи. Довольно многие паразиты занимают по своей специализации к растениям-хозяевам промежуточное положение. У них проявляется заметная тенденция к узкой специализации и в тоже время сохраняется способность поражать довольно много видов и родов растений-хозяев.
У видов паразитов, для которых узкая специализация не доказана, разновидности или специализированные формы обычно не выделяют, но определение рас для них возможно (например, у Phytophthora megasperma и P. infestans).
Ph. megasperma способна поражать растения из семейств Бобовых (люцерна, соя, клевер), Розовых (яблоня, груша), Крестоцветных (цветная капуста) и Ивовых (тополь). Однако исследования, сделанные с привлечением морфологических (строение оогония) и молекулярных (митохондриальная ДНК и фрагменты рестрик-ционного полиморфизма ДНК) маркеров, позволили выделить разновидности Ph. megasperma var. megasperma (поражает различные растения-хозяева) и Ph. megasperma var. sojae (поражает растения-хозяева преимущественно из семейства Бобовых). У Ph. megasperma var. sojae выделили f. sp. glycinea (поражает сою) и f. sp. medicaginis (поражает люцерну, а также нут и эспарцет). Наконец, последние исследования выявили настолько существенные различия между этими разновидностями и специализированными формами, что они получили статус отдельных видов Ph. megasperma, Ph. sojae и Ph. medicaginis. Этот случай, с одной стороны, показывает тенденцию к узкой специализации у исходно широкоспециализированного вида, с другой стороны, здесь можно видеть сохранение в значительной степени комплексной структуры у внутривидовых группировок.
Phytophthora infestans поражает только картофель и томат, реже баклажаны и дикие виды из семейства Пасленовых. Предполагали специализацию Ph. infestans по отношению к основным родам растений-хозяев — картофелю и томату. Раса ТО заражала преимущественно картофель, расы Т1 и Т2 — в основном томат. Однако исследования 1990-х годов в ряде случаев выявили комплексное распределение этих рас, не всегда приуроченное к картофелю или томату. Отмечено, что штаммы Ph. infestans с томата активно поражают и томат, и картофель, большинство штаммов с картофеля (но не все) значительно лучше поражают картофель, чем томат. Таким образом, имеется лишь ограниченная приуроченность штаммов паразита к определенному роду растения-хозяина. Сходная ситуация сложилась у Ph. palmivora, часть штаммов которой лучше поражала гевею, чем какао/другая часть — какао, чем гевею.
Erysiphe communis вызывает мучнистую росу у растений очень многих семейств (например, Бобовых и Пасленовых). В целом, этот вид может характеризоваться очень широкой специализацией, однако по мере его изучения обнаруживают много специализированных форм, приуроченных к определенным семействам, родам и видам растений-хозяев. Er. cichoracearum также вызывает мучнистую росу у растений из многих семейств. Ядро этого вида связано с родами из семейства Сложноцветных, однако Ег. cichoracearum f. sp. cucurbitacearum поражает растения из семейства Тыквенных (особенно огурец, дыню и тыкву).
Цветковое растение-паразит Orobanche foetida (заразиха вонючая) паразитирует на только трех родах из семейства Бобовых — чине, лядвенце и эспарцете. Другие виды заразих могут паразитировать как на одном виде растения-хозяина, так и на растениях-хозяевах из разных семейств.
Таким образом, многие виды паразитов имеют промежуточную специализацию. Это связано с микроэволюционными процессами в их популяциях, которые, вероятно, еще не завершились. Во всех случаях имела место дивергенция исходных комплексных групп паразитов, направленная на совершенствование паразитизма по отношению к определенным видам растений-хозяев. Однако надо признать, что для некоторых случаев (как, например, с комплексом Phytophthora megasperma) ситуация изучена не полностью и до сих пор вызывает у специалистов много споров.
Физиологические расы и методы их определения.Расы паразитов поражают определенные сорта растений-хозяев. Расы ржавчинного гриба Puccinia graminis были открыты американским ученым Стэкменом, расы Phytophthora infestans — шотландским селеционе-ром Блэком в 1950—1960-е годы.
Понятие «физиологические» связано с тем, что способность рас преодолевать устойчивость одних сортов и неспособность преодолевать устойчивость других связана с различными физиологическими механизмами. Эти различия связаны с продуктами обмена веществ паразита и в клетках растения-хозяина.
Физиологические расы известны для патогенных бактерий, грибов и цветковых растений-паразитов. У вирусов различают штаммы, отличающиеся друг от друга по структуре ДНК и патогенности по отношению к растениям-хозяевам.
Для определения рас используют растения-дифференциаторы. Они относятся к сортам и гибридам, характеризующимся определенной вертикальной устойчивостью по отношению к возбудителю болезни. В расах присутствуют факторы вирулентности, в дифференциаторах — факторы вертикальной устойчивости. Между расой паразита и сортом растения-хозяина происходит дифференциальное (избирательное) взаимодействие. Оно заключается в следующем.
Если та или иная раса паразита преодолевает устойчивость сорта, то наблюдается реакция совместимости (цв. вклейка, рис. 10). На пораженных органах проявляются основные симптомы (например, спороношение Phytophthora infestans или пустулы Puccinia graminis). Если раса паразита сталкивается с устойчивостью сорта, то на органах растения наблюдается реакция сверхчувствительности (цв.вклейка, рис. 11) — основной механизм активного иммунитета растений. Она проявляется в виде мелких некрозов в местах внедрения спор возбудителя в растение-хозяина.
Авирулентная раса паразита (без факторов вирулентности) способна поражать только восприимчивые сорта (без факторов вертикальной устойчивости). Устойчивые сорта авирулентная раса, как правило, не поражает.
Вирулентная раса паразита (с фактором или факторами вирулентности) способна поразить восприимчивые сорта. Преодолеть устойчивость сорта с фактором или факторами вертикальной устойчивости вирулентная раса сможет в том случае, если они будут полностью соответствовать фактору или факторам вертикальной устойчивости. Если этого соответствия не будет хотя бы по одному из факторов, то раса паразита не сможет преодолеть устойчивость сорта.
Генетически факторы вирулентности определяются vir-генами (virulence — вирулентность), а факторы вертикальной устойчивости — R-генами (resistance — устойчивость). В ряде исследований на основе генетического анализа было показано, что один фактор (вирулентности или вертикальной устойчивости) определяется одним геном (vir- или R-геном соответственно) с полным доминированием. Вирулентность — признак рецессивный, а вертикальная устойчивость — доминантный. В последние годы посредством ПЦР-анализа ведется уточнение локусов (последовательностей ДНК), соответствующих тому или иному vir-гену паразита или R-гену растения-хозяина.
Для выявления рас Phytophthora infestans существует набор растений-дифференциаторов Блэка. Он состоит из сортов и гибридов с определенными R-генами: R1...R11. Набор Блэка признан селекционерами всего мира. Он хранится и поддерживается в Международном центре по картофелю (IPC) в Перу. При определении рас растения-дифференциаторы (или листья с них, разложенные во влажные камеры) заражают суспензией конидий изолятов Ph. infestans. Заражение проводят в помещении с низкой освещенностью и высокой относительной влажностью, то есть в контролируемых условиях.
Дифференциатор R1 способен выявить две расы Ph. infestans, R1 и R2 — четыре, Rl, R2 и R3 — восемь рас и т. д. (табл. 6).

Чем больше растений-дифференциаторов растения-хозяина используют, тем больше рас можно выявить. Общее число возможных рас (7V) определяется формулой