Характеристика отдельных видов и групп бактерий рода Pseudomonas
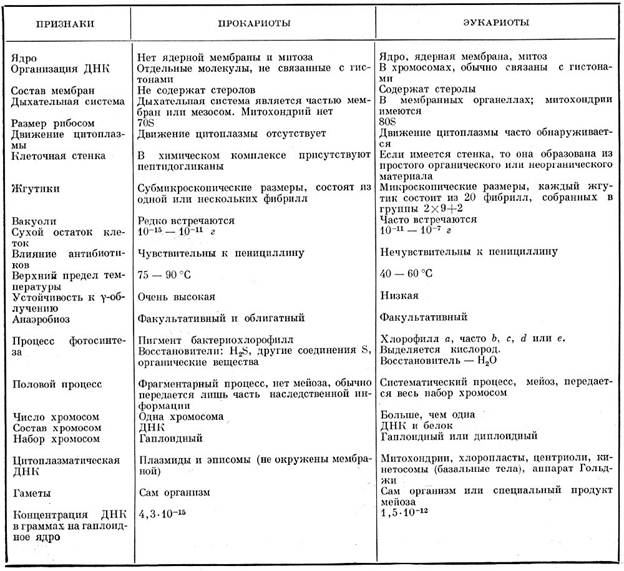
Таблица. Сопоставление признаков прокариотных и эукариотных организмов
Таблица. Основные систематические группы
Основные систематические группы бактерий и актиномицетов
В настоящей книге авторы придерживались в основном системы бактерий и актиномицетов, разработанной крупнейшим советским микробиологом Н. А. Красильниковым в его «Определителе бактерий и актиномицетов» (1949).
Эубактерии и родственные им организмы, по Н. А. Красильникову, подразделяются на 4 класса:
Eubacteriae (истинные бактерии). Actinomycetes (актиномицеты). Myxobacteriae (миксобактерии). Spirochaetae (спирохеты).В класс Eubacteriae (истинные бактерии) входят организмы с неветвящимися клетками и прочными клеточными стенками, имеющими форму мелких тонких палочек, реже кокков, находятся также нитевидные и спиралевидные формы. Часть эубактерий обладает подвижностью, часть — неподвижные. Движение осуществляется при помощи жгутиков простого строения, состоящих из взаимно перевитых фибрилл.
Часть эубактерий способна формировать эндогенно (внутри клеток) термоустойчивые споры, большинство же представлено неспороносными грамотрицательными формами. Размножаются эубактерии делением, некоторые нитчатые бактерии образуют специальные репродуктивные клетки — гонидии.
Класс Eubacteriae объединяет порядки: Eubacteriales (эубактерии), Chlamydobacteriales (хламидобактерии), Ferribacteriales (железобактерии), Thiobacteriales (серобактерии).
В пределах этого класса могут рассматриваться на уровне порядков почкующиеся бактерии (Hyphomicrobiales), приводящиеся в «Определителе бактерий и актиномицетов» Н. А. Красильникова в группе неклассифицированных организмов, а также многоклеточные бесцветные бактерии, сходные с сине-зелеными водорослями и выделяемые М. А. Пешковым в отдельный порядок Caryophanales.
| EUBACTERIALES. Одноклеточные бактерии шаровидной, палочковидной и спиралевидной формы. Грамотрицательные и грамположительные. Раэммножаются поперечным делением клеток. Большинство подвижные. Жгутики расположены полярно на концах клеток или перитрихиально. Некоторые представители способны образовывать эндогенно термоустойчивые споры. Разнообразны по физиологическим свойствам: автотрофы и гетеротрофы. Очень широко распространены в природе. | 1. Грамотрицательные неспоробразующие палочки с одним жгутиком. Типичные представители порядка. Сильно различаются по физиологическим признакам: гетеротрофизм, паразитизм, облигатная галофилия. ) | Pseudomonas, Xanthomonas, Azotomonas, Acetomonas, Aeromonas, Zymomonas, Acetobacter, Bdellovibrio, Halobacterium |
| 2. Неспороносные грамотрицательные палочки с перитрихиальнм жгутикованием. Некоторые представители неподвижны. Гетеротрофы. Есть патогенные формы возбудители тифа, дизентирии, чумы. Представители родов Azotobacter, Bejerinckia могут фиксировать атмосферный азот. Некоторые виды размножаются почкованием (Agrobacterium, Tuberoidobacter). Имеются организмы необычной морфологии: Seliberia, Tuberoidobacter | Bacterium, Serratia, Salmonella, Shigella, Erwinia, Aerobacter, Klebsiella, Proteus, Pasteurella, Chromobacterium, Flavobacterium, Fusobacterium, Achromobacter, Alcaligenes, Beneckea, Agarbacterium, Azotobacter, Bejerinckia, Rhizobium, Agrobacterium, Tuberoidobacter, Seliberia, Haemophilus, Bordetella, Moraxella | |
| 3. Bacillaceae. Грамположительные и грамотрицательные палочковидные бактерии. Способны формировать термоустойчивые эндоспоры. Аэробы (род Bacillus) и анаэробы (роды С1оstridium, Desulfotomaculum). Гетеротрофы | Bacillus, Clostridium, Desulfotomaculum | |
| 4. Spirillaceae. Грамотрицательные бактерии. Клетки имеют вид слабо (вибрионы) или сильно (спириллы) изогнутых палочек с жесткой клеточной стейкой. Аэробы и анаэробы. Гетеротрофы. Есть возбудители болезней (холера) | Vibrio, Cellvibrio, Cellfalcicola, Microcyclus, Spirosoma, Benobacter, Spirillum, Desulfovibrio, Selenomonas | |
| 5. Planococcaceae. Шаровидные Грамположительные подвижные бактерии. Клетки одиночные (род Р1аnococcus) или соединены в группы (род Planosarcina). Гетеротрофы | Planococcus, Planosarcina | |
| FERRIBACTERIALES. Сборная группа одноклеточных бактерий, обладающих способностью откладывать вокруг клеток соединения железа | 1. Ferribacteriaceae. Клетки образуют слизистые стебельки, на которых откладывается гидрат окиси железа. Гетеротрофы | Gallionella, Nevskia, Pasteuria |
| 2. Siderocapsaceae. Клетки образуют слизистую капсулу, иногда капсул нет. Гетеротрофы | Siderocapsa, Sideromonas, Ochrobium, Naumanniella, Siderococcus, Ferrobacillus, Ferribacterium | |
| HYPHOMICROBIALES. Гетеротрофные грамотрицательные неспорообразующие бактерии, клетки которых обладают протоплазматическими выростами - гифами. Размножаются путем образования почек на концах гиф. | Hyphomicrobiaceae. Характеристика та же, что и для порядка | Hyphomicrobium, Pedomicrobium, Ancalomicrobium, Prosthecomicrobium, Blastobacter, Blastocaulis |
| THIOBACTERIALES. Сборная группа одноклеточных бактерий, которые при окислении сероводорода откладывают внутри клеток капельки серы | Achromatiaceae. Типичные представители порядка. Бесцветные одноклеточные бактерии. Аэробы. Гетеротрофы | Achromatium, Thiophysa, Thiosphaerella, Thiovulum |
| CHLAMYDOBACTERIALES. Нитчатые бактерии. Гетеротрофы. Многие способны откладывать в слизистых чехлах (влагалищах) гидраты окиси железа. Растут в виде нитей. Молодые клетки, выходящие из чехла, подвижны, обладают полярным и субтерминальным жгутиками | Chlamydobacteriaceae. Растут в виде нитей, покрытых слизистой капсулой. Гетеротрофы. Аэробы | Sphaerotilus, Leptotrix, Crenotrix, Thiothrix |
| CARYOPHANALES. Многоклеточные нитчатые (трихомные) бактерии. Сходны с синезелеными водорослями (цианобактериями), но бесцветны. Нити подвижные и неподвижные. Некоторые виды образуют эндогенные споры | 1. Oscillospiraceae. Образуют споры. Трихомы различной длины, состоят из дисковидных клеток. Подвижны, жгутпкование перитрихиальное. Анаэробы | Oscillospira |
| 2. Caryophanaceae. Спор не образуют. Трихомы часто подвижны | Caryophanon | |
| 3. Arthromitaceae. Трихомы состоят из цилиндрических клеток. Неподвижны. Споры образуются на дисталъных концах трихомов | Arthromitus, Simonsiella | |
| 4. Pontotricaceae. Трихомы неподвижны, состоят из цилиндрических клеток. Размножаются гормогониями и одноклеточными гонидиями | Pontothrix | |
| ACTINOMYCETALES. Прокариотные микроорганизмы, клетки которых способны к ветвлению и образованию мицелия. Подвижных клеток не образуют. Очень широко распространены в природе | 1. Actinomycetaceae. Типичные представители актиномицетов с хорошо развитым мицелием. Споры формируются путем одновременного расчленения спороносцев на длинные цепочки спор | Actinomyces, Proactinomyces, Chainia, Actmopicnidium |
| 2. Micromonosporaceae. Актиномицеты с хорошо развитым мицелием. Споры формируются путем последовательного отчленения кончика спороносна. Число спор небольшое | Micromonospora, Microbispora, Micropolyspora, Actinobifida, Promicromonospora | |
| 3. Streptosporangiaceae. Актиномицеты с хорошо развитым мицелием. Образуют спорангии, в которых формируются множественные споры | Streptosporangium, Micrcsporangium, Amorphosporangium, Elytrosporangium, Actinosporangium, Intrasporangium | |
| MYCOBACTERIALES. Бактерии, не образующие мицелия. Клетки палочковидные, способны ветвиться. Спор не образуют. Размножаются простым делением или дроблением | Mycobacteriaceae Характеристика та же, что и для порядка | Mycobacterium, Mycococcus, Propionibacterium, Lactobacterium, Pseudobacterium |
| COCCALES. Бактерии с кокковидпыми клетками различных размеров и неправильной конфигурацией. Неподвижные. Спор не образуют. Размножаются делением, перешнуровыванием и почкованием | Соссасеае. Характеристика та же, что и для порядка | Micrococcus, Nitrosococcus, Diplococcus, Neisseria, Streptococcus, Sarcina, Peptostreptococcus, Pediococcus, Leuconostoc, Staphylococcus, Gaffkya, Methanococcus, Peptococcus, Veillonella, Aerococcus |
| ACTINOPLANALES. Прокариотные микроорганизмы, клетки которых способны ветвиться и образовывать мицелий. В процессе развития образуются подвижные клетки - споры или фрагменты нитей мицелия | 1. Actinoplanaceae. Актиномицеты, образующие хорошо оформленные спорангии. Внутри спорангиев формируются подвижные споры | Actinoplanes, Spirillospora, Ampullariella, Pilimelia, Kitasatoa |
| 2. Dermatophilaceae. Актиномицеты, формирующие подвижные споры путем дробления нитей мицелия. Образуемые скопления спор имеют неопределенную форму, без резких очертаний | Dermatophilus, Proactinoplanes | |
| 3. Planosporaceae. Актиномицеты, формирующие подвижные споры на коротких спороносцах воздушного мицелия. Споры покрываются слизистой капсулой, имитирующей оболочку сумки. Однако спорангии, как таковые, отсутствуют | Planospora, Planobispora, Sporichtia, Dactylosporangium | |
| MYXOBACTERIALES. Грамотрицательные бактерии. Клетки гибкие, способны к скользящему движению в слизи. Многие образуют слизистые колонии и плодовые тела | 1. Promyxobaeteriaceae. Не образуют плодовых тел. Образуют либо не образуют микроцист. Активно разлагают клетчатку | Cytopliaga, Sporocytophaga, Flexibacter |
| 2. Archangiaceae. Колонии не имеют правильной формы. Нет оформленных цист | Archangium, Stelangium | |
| 3. Sorangiaceae. Цисты угловатые, собраны в кучу, окруженную общей оболочкой | Sorangium | |
| 4. Polyangiaceae. Оформленные цисты с четкими оболочками. Плодовые тела пигментированы, ветвятся | Polyangium, Synangium, Меlittangium, Podangium, Chondromyces | |
| 5. Myxococcaceae. Палочковидные клетки укорачиваются в цистах, образуют споры | Myxococcus, Chondrococcus, Angiococcus | |
| SPIROCHAETALES. Клетки спирально извиты, змеевидно подвижны, обладают осевой плотной нитью, вокруг которой закручена клетка | Spirochaetaceae. Включает 6 родов, различающихся морфологически, по размерам клетки (мелкиклетки Spirochaeta, гигантские - Cristispiга). Есть паразиты (Borrelia) | Spirochaeta, Saprospira, Cristispira, Borrelia, Treponema, Leptospira |
| MYCOPLASMATALES. Клетки разнородны по форме, размерам, не имеют клеточных стенок. Гетеротрофы. Паразиты и сапрофиты | 1. Mycoplasmataceae. Микоплазмы, не нуждающиеся в стерине | Mycoplasma, Thermomycoplasma |
| 2. Acholeplasmataceae. Стеринзавпсимые микоплазмы | Acholeplasma | |
| Rickettsiales. Мелкие палочковидные и кокковидные грамотрицательные бактерии. Не растут на средах. Облигатные внутриклеточные паразиты, возбудители болезней человека и животных. Аэробы, содержат цитохромы. | 1. Rickettsiaceae. Типичные представители порядка. Паразитируют в клетках тканей животных и растений | Rickettsia, Dermacentroxenus, Coxiella |
| 2. Ehrlichiaceae. Паразитируют в лейкоцитах | Ehrlichia, Cytoecetes | |
| 3. Bartonellaceae. Паразитируют в эритроцитах | Bartouella, Eperythrozoon, Haemobartolpnella, Grahamella | |
| CHLAMYDIALES. Облигатные энергетические паразиты, не способные генерировать АТФ. Анаэробы, не содержат цитохромов. Характеризуются сложным циклом развития | Chlamydiaceae. Характеристика та же, что и для порядка | Chlamydia |
Класс Actinomycetes (актиномицеты) состоит из организмов, обладающих мицелиальными, нитевидными, палочковидными и кокковидны-ми грамположительными клетками. В отличие от бактерий, входящих в другие классы, для типичных представителей актиномицетов (высшие формы) характерно наличие хорошо выраженного ветвящегося септированного или не-септированного мицелия.
Имеются и переходные к типичным актиноми-цетам низшие формы — микобактерии, мико-кокки и микрококки, для которых характерны ветвление на ранних стадиях развития, образование боковых выростов — рудиментов веток.
Класс актиномицетов подразделяется на порядки: Actinomycetales (актиномицеты, не образующие подвижных клеток), Actinoplanales (актиномицеты с подвижными клетками), Мусоbacteriales (микобактерии) и Coccales (кокки).
Класс Myxobacteriae (миксобактерии). Организмы этого класса представлены палочковидными и кокковидными клетками, обладающими, тонкой эластичной (неригидной) клеточной стенкой, благодаря чему они при движении могут изменять свою форму, изгибаться. Жгутиками не обладают. Движение осуществляется по типу скольжения. Миксобактерии способны образовывать микроцисты, а также особые плодовые тела, имеющие самую различную форму.
В класс миксобактерии входит один порядок — Myxobacteriales.
Порядок эубактерии (EUBACTERIALES)
В строгом смысле к истинным бактериям относят одноклеточных бактерий с ригидными клеточными стенками, гетеротрофов, перемещающихся (если обладают подвижностью) с помощью жгутиков. В эту группу входит преобладающее количество прокариотных микроорганизмов.
Клетки эубактерий очень малы и вместе с тем сильно различаются по размерам: от карликовых форм (диаметром около 0,125 мкм) до гигантов, длиной до 10 мкм (Ghromatium okenii). В природных субстратах (илах, почвах) можно встретить стебельковых бактерий, достигающих в длину вместе со стебельком 100 мкм и более. Следовательно, гиганты превышают карликов по размерам в 100 и 1000 раз. Большинство палочковидных бактерий по длине не превышают 5 мкм, по толщине 1 мкм. У многих псевдомонад диаметр клетки равен 0,4—0,7 мкм, у ряда почвенных форм — 0,2—0,3 мкм.
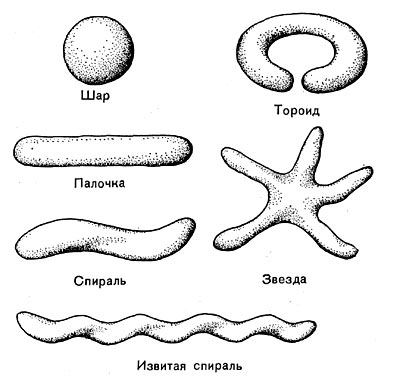
Рис. 14. Основные формы бактериальных клеток. Схема.
Разнообразие форм бактериальных клеток невелико. Основные формы бактерий — это шары (кокки), палочки (прямые, изогнутые или извитые), тороиды и звезды (рис. 14). Клетки многих видов неспорообразующих и спорооб-разующих бактерий выглядят как палочки. Спириллы имеют форму спирали, и ряд близких видов — форму тороида.
Поверхность клеток бактерий имеет часто придаточные структуры. Наиболее распространенными являются жгутики и ворсинки (фимбрии, нитевидные придатки клеток).
Фимбрии (или ворсинки) короче и тоньше жгутиков, имеют вид жестких прямых нитей и, покрывая всю клетку, создают ворсистую поверхность (рис. 15, 16). Длина фимбрий от 0,3—4 мкм, ширина — 0,01 мкм. Иногда они сливаются в комки, придавая неопрятный вид клетке; в других случаях поверхность клеток покрыта войлокообразным чехлом, состоящим из сплетений тонких нитей. Наряду с фимбриями можно видеть капсульный материал, как бы застрявший между нитями фимбрий, выполняющих роль арматуры. Нередко можно встретить лентовидные придатки у клеток. В среде обитания фимбрий выполняют прикрепительные функции, обеспечивая прикрепление клеток к частицам почвы, органических остатков, объединение клеток в группы (рис. 16). Есть сведения о том, что по коротким нитям, соединяющим две клетки и имеющим внутри канал, происходит обмен наследственной информацией. Среди побочных функций фимбрий — защита клетки от нападения паразитов, так как успех атаки паразитов обеспечивается непосредственным контактом с поверхностью клетки хозяина.
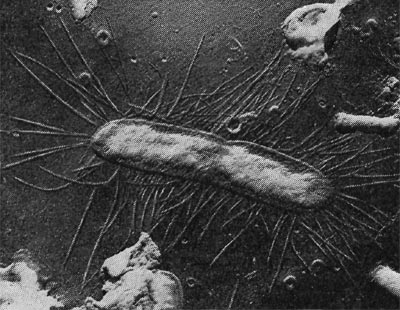
Рис. 15. Палочковидная бактерия с фимбриями. Увел. X 15 000.
Изучение химической природы ворсинок встретилось со значительными трудностями, так как потребовалось их отделить от клетки, «побрить» клетку. К счастью, для исследователей это оказалось возможным проделать в дезинтеграторах — приборах, разрушающих клетки. При этом ворсинки обрываются, после чего их отделяют на центрифугах и накапливают. Как оказалось, они состоят из белка, обладают гидрофобностью (водоотталкивающими свойствами) и локализованы на клеточной стенке, представляя собой как бы «взлохмаченную» стенку клеток.
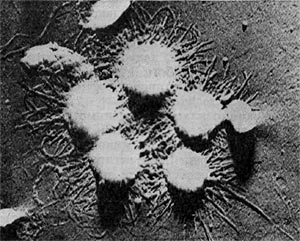
Рис. 16. Кокки с фимбриями. Увел. X 12000.
Другим типом придатков, свойственным многим бактериям, являются жгутики. По структуре они представляют собой гомогенную белковую нить или плотно свернутую косу из 2—3 нитей. Длина жгутиков 3—12 мкм, толщина — 0,02 мкм. Жгутики волнообразно изогнуты и могут быть расположены по всей поверхности клетки (перитрихиально) либо на полюсах (монотрихиально). Они составляют до 2% сухой массы бактерии. Жгутики являются органом движения бактерий, хотя известны случаи существования бездействующих (парализованных) жгутиков. Прикрепление жгутиков к клетке связано с цитоплазматической мембраной (рис. 17, 18). Основание жгутика прикреплено к специальной мембране. При делении у монотрихов дочерняя клетка приобретает полярный жгутик в том месте, где завершилось деление (рис. 19,7). В случае перитри-хиального жгутикования новые и ранее существовавшие жгутики распределяются равномерно (рис. 19,2).
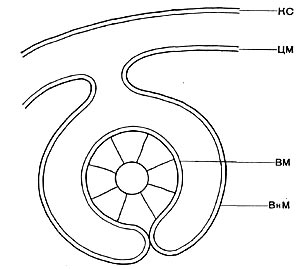
Рис. 17. Схема структуры зоны прикрепления жгутика в клетке бактерии: КС — клеточная стенка; ЦМ — цитоплазматическая мембрана; ВМ — внутренняя мембрана прикрепительного тельца жгутика; ВнМ — внешняя мембрана прикрепительного тельца (по Таушель и др.).
У многих микроорганизмов жесткая клеточная стенка окружена слоями слизистого материала, образующего капсулы разной плотности и толщины (рис. 20). Основными компонентами капсул являются вода и полимеры — полисахариды. Особенно обильное образование слизи наблюдается у многих микроорганизмов в тех случаях, когда среда содержит сахарозу. Молочнокислая бактерия Leuconostoc mesen-teroides за несколько часов превращает раствор в студень — декстран, который используют как заменитель крови, для повышения вязкости водных растворов. Препарат из декстрана — сефадекс — применяется в лабораториях в качестве «молекулярного сита» для разделения веществ с большой молекулярной массой.
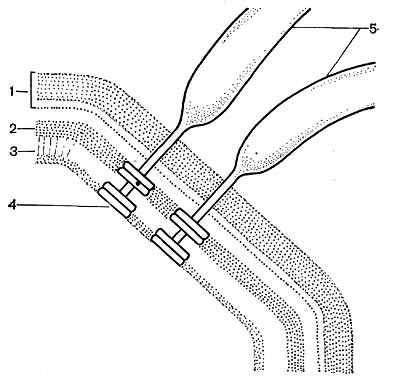
Рис. 18. Схема прикрепления жгутика: 1 — клеточная стенка; 2 — цитоплазматическая мембрана; 3 — мембрана жгутиков; 4 — диски основания жгутиков; 5 — жгутики (по Броку, 1970).
Основным структурным компонентом клетки бактерий является клеточная стенка, которая придает протопласту микроба характерную для него форму. Когда целостность стенки нарушается, образуется сферический протопласт (при участии внутриклеточного осмотического давления, достигающего 20—50 атм и более). Клеточная стенка составляет от 10 до 50% сухой массы бактерии. Количество материала клеточной стенки увеличивается с возрастом.
Исследования последнего десятилетия по биохимии клеточной стенки бактерий являются крупнейшими достижениями современной микробиологии, позволившими понять молекулярную организацию микробной стенки. Как уже упоминалось ранее, в зависимости от строения клеточной стенки эубактерии разделяются на 2 группы — грамположительные и грамотрицательные. В стенках грамположи-тельных бактерий очень высокое содержание гликопептидов (95% от суммы веществ стенки), однако их строение более простое, чем стенок грамотрицательных бактерий. В стенках грам-отрицательных бактерий лишь 5% гликопептидов. Этот слой покрыт рыхлым слоем белка, который, в свою очередь, прикрыт липополиса-харидным и липопротеидным слоями, пронизанными каналами (рис. 21 и 6).

Рис. 19. Распределение и образование жгутиков во время деления клеток: 1 — организм с полярным жгутиком, 2 — перитрих. Схема.
Муреиновый мешок выполняет у бактерий роль опорного каркаса. У грамположительных бактерий он многослоен, у грамотрицательных — однослоен. По существу дела, этот слой представляет собой одну молекулу, покрывающую бактериальную клетку.
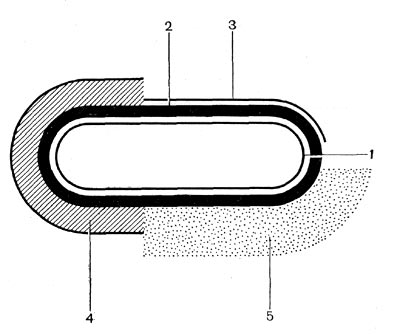
Рис. 20. Схема взаиморасположения внешних слоев клетки бактерий: 1 — цитоплазматическая мембрана, 2 — клеточная стенка, 3 — микрокапсула, 4 — капсула, 5 — слизистый слой.
Все описанные здесь особенности прокари-отных организмов являются исключительно своеобразными, свойственными представителям этого удивительного мира микроскопических существ. Внешний вид и функционально аналогичные органы (жгутики, реснички и др.) оказываются иными у эукариотных клеток.
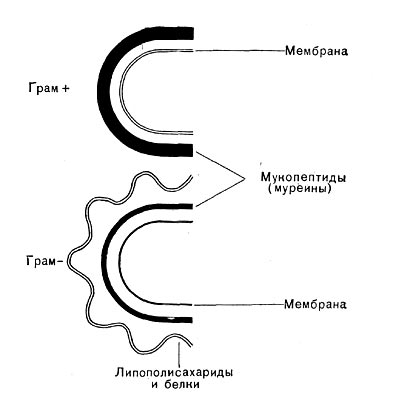
Рис. 21. Схема строения клеточных стенок грамположительных (Грам+) и грамотрицательных (Грам -) — бактерий.
Уникальны также структура и состав клеточных стенок бактерий. Структурные элементы стенок представляют ахиллесову пяту бактерий, так как некоторые лекарственные препараты действуют только на стенки бактерий, не затрагивая эукариотных клеток высших организмов.
Многие победы современной медицины основаны на специфичности клеточных стенок наших друзей и врагов — прокариотов.
Большие различия между эукариотами и прокариотами обнаруживаются и в мембранном аппарате клеток (в химическом составе и структуре). Так, например, у бактерий нет истинных митохондрий, играющих роль энергетических депо клеток. Их функции выполняют простые мембранные структуры (мезосомы). Особенности питания, транспорт веществ через стенку, богатейший спектр веществ, синтезируемых бактериями, особенности дифференцировки клеток, большая скорость размножения и адаптации к среде делают прокариотов истинными хозяевами и вершителями процессов круговорота веществ в биосфере.
Сопоставление некоторых свойств прокариотных и эукариотных организмов приведено в таблице на стр. 208.
Семейство псевдомонады (PSEUDOMONADACEAE)
Типичными представителями эубактерии являются грамотрицательные неспороносные бактерии, объединяемые в семейство Pseudomo-nadaceae. Название семейства происходит от двух греческих корней: «псевдо» — сходный и «монас» — название группы простейших (животных) с полярно расположенными жгутиками. Поэтому к псевдомонадам относят как палочковидные бактерии с полярно расположенным жгутиком, так и слабоизогнутые палочки, физиологически крайне специализированные ав-тотрофные хемосинтезирующие бактерии (Нуdrogenomonas, Nitrosomonas, Thiobacillus) и обычные гетеротрофные бактерии (Pseudomonas), т. е. смешиваются представители питания — автотрофного и гетеротрофного.
Следовательно, в семейство Pseudomonada-сеае входят грамотрицательные палочки, растущие в аэробных условиях или получающие энергию за счет анаэробного дыхания либо за счет связанного кислорода (денитрификация), но не брожения. Псевдомонады используют широкий спектр органических веществ, в том числе циклических соединений (гетероциклических и ароматических). Благодаря «всеядности» псевдомонады распространены повсеместно: в воде, почве, илах, переносятся с током воздуха. Многие из них образуют водорастворимые и флюоресцирующие пигменты.
Патогенность у псевдомонад выражена слабо, но даже такой обычно хорошо знакомый микробиологам сапрофит, как Pseudomonas aerugi-nosa, часто является возбудителем вторичной инфекции долго не заживающих ран и язв у человека и животных. Пигменты, выделяемые этим микробом, придают гною сине-зеленую окраску. Вспышки энтерита у новорожденных вызываются этой бактерией. Она же вызывает заболевания растений (табак и салат) и смертельные заболевания домашней птицы. Представители другого рода (объединяет 65 видов) — Xanthomonas — наносят большой вред сельскому хозяйству, деревообрабатывающей и другим видам промышленности, где используется растительное сырье. Они вырабатывают ферменты, которые разрушают протопектины — вещества, цементирующие клетки растений. Этот процесс приводит к ослизнению растительной массы, а растение — к гибели.
Род псевдомодас (PSEUDOMONAS)
Бактерии этого рода широко используются в хозяйственной практике, а также в качестве моделей для многочисленных теоретических исследований, поэтому мы и остановимся на этих микроорганизмах подробнее.
В 50—60-х годах нашего столетия в мировой и отечественной литературе появилось большое количество научных исследований о бактериях рода Pseudomonas, в частности об их флуоресцирующей группе. Эти исследования касаются вопросов использования флуоресцирующих бактерий для решения ряда практических и теоретических задач. Существует много патентов и научно-исследовательских работ американских, японских и советских авторов, где подробно представлены характеристики продуцентов, состав сред для выращивания бактерий и описаны условия культивирования для получения значительных количеств различных биологически активных соединений.
Так, при использовании в качестве продуцентов некоторых штаммов бактерий Ps. fluorescens осуществляется биосинтез органических кислот: глюконовой, 2-кетоглюконовой, а-кетоглутаровой и пировиноградной. Для получения аминокислот: глутаминовой, l - аспарагиновой, валина и метионина — также используются различные представители бактерий Pseudomonas fluorescens и Ps. aeruginosa.
Из окрашенных веществ, синтезируемых бактериями рода Pseudomonas, были выделены химические вещества, обладающие антибиотическими свойствами, — пиоцианин, хлоро-рафин, оксихлорорафин, феназин-1-карбоновая кислота и эругинозин. Все перечисленные пигменты обладают антибиотической активностью против грамположительных и грамотрицатель-ных бактерий, дрожжей и грибов.
Кроме антибиотиков, в состав окрашенных веществ, синтезируемых псевдомонадами, входят витамины. Советские и зарубежные исследователи еще в 30-х годах нашего столетия отмечали, что в состав желто-зеленого флуоресцирующего пигмента, синтезируемого псевдомонадами, входят рибофлавин, фолиевая кислота, птерин. Использование современных методов исследования позволило выделить в кристаллически чистом виде и определить составные части витаминов птеридинового и каротиноид-ного типов, образуемых псевдомонадами.
В работах советских и зарубежных исследователей последнего десятилетия отмечается, что флуоресцирующие псевдомонады могут быть продуцентами ферментов. Так, советские исследователи изучают процесс синтеза фермента аспарагиназы, используя Ps. fluorescens. Ac-парагиназа обладает канцеростатическими действиями. В ФРГ организовано производство фермента пероксидазы. В качестве продуцента пероксидазы также используют бактерии Ps. fluorescens.
В практике контроля пищевых продуктов флуоресцирующие псевдомонады издавна известны как индикаторы на зараженность.
Интенсивное развитие бактерий на пищевых продуктах сопровождается выделением флуоресцирующего пигмента. Этот признак применяется при отбраковке пищевых продуктов.
Решение ряда теоретических вопросов осуществляется с помощью модельных опытов с бактериями рода Pseudomonas: изучаются некоторые пути углеводного и аминокислотного обмена, отдельные этапы процесса дыхания микроорганизмов.
Японские и американские исследователи использовали псевдомонады для получения кристаллического цитохрома и изучения его аминокислотного состава.
Бактерии рода Pseudomonas широко распространены в природе. Их можно встретить в воздухе, почве, морских и пресных водоемах, сточных водах и иле, нефти и на газовых месторождениях. Псевдомонады были обнаружены на пищевых продуктах, телах животных, растениях, а также в гнойных ранах и экскрементах больных млекопитающих.
Интерес к этой группе бактерий все больше возрастает. Кроме микробиологов, фитопатологов и медицинских бактериологов, псевдомонады привлекают внимание также и ученых других профилей: лимнологов, почвоведов и т. д.
По Красильникову, бактерии рода Pseudomonas монолитны по морфологическим и очень разнообразны по культуральным и физиологическим признакам.
Клетки псевдомонад представляют собой мелкие одиночные грамотрицательные палочки. Спор и выростов не образуют, подвижны, имеют полярно расположенные жгутики. Число жгутиков у разных видов колеблется. Клетки в культурах часто объединяются в небольшие комочки или зерна, окруженные толстой слизистой оболочкой, иногда резко очерченной, — зооглеи.
К настоящему времени накопилось достаточно сведений, чтобы использовать некоторые биохимические признаки для таксономической характеристики рода. Так, например, химический состав клеточной стенки бактерий постоянен и, как правило, не изменяется на различных стадиях развития культуры или при возникновении мутаций. У бактерий рода Pseudomonas процент Г + Ц (гуанин + цитозин) в ДНК колеблется в пределах 58—69%.
Колонии бактерий очень разнообразны: слизистые и пастообразные, выпуклые и плоские, крупные и мелкие. У многих видов отмечается внутренняя структура колоний. Если их рассматривать в микроскопе при малом увеличении, то в одних случаях можно обнаружить мелкозернистую колонию; в других — ячеистую, напоминающую соты; в третьих — колонии в виде мелких комочков или зерен. Большинство видов имеет колонии без внутренней структуры — под микроскопом они выглядят как однородная гомогенная масса. Псевдомонады хорошо растут на обычных питательных средах — сложных органических.
Культуры различаются между собой способностью разлагать белки, использовать углевод, расщеплять крахмал, клетчатку, углеводороды, соединения ароматического ряда и другие сложные по составу вещества.
Большинство бактерий рода Pseudomonas обладает гетеротрофным типом обмена веществ, т. е. для построения тела им требуется готовое органическое вещество. Биосинтетические процессы, при этом осуществляются за счет обмена окислительного типа, где кислород является конечным акцептором электронов, перенос которых связан с системой цитохромов. Некоторые представители этого рода могут существовать за счет анаэробного нитратного дыхания, другие используют энергию окисления водорода. Многие виды псевдомонад образуют пигменты, различные по окраске и химической природе; некоторые синтезируют витамины, антибиотики, токсины.
Среди представителей бактерий рода Pseudomonas есть формы, токсичные для животных организмов (патогенные бактерии). Имеется также немало фитопатогенных видов, поражающих растения.
В «Определителе бактерий и актиномицетов» (1949) Н. А. Красильникова описано более 200 видов бактерий рода Pseudomonas. Это наиболее полное и пока единственное систематическое описание, где представлены характеристики почти всех известных псевдомонад, которые распределены между собой в зависимости от характера своих признаков.
Для разделения этих бактерий на виды использован легко наблюдаемый признак пигментации, а также морфологические, культу-ральные и некоторые физиологические свойства. Особенно отмечен признак токсичности бактерий по отношению к животным и растительным организмам.
Ps. aeruginosa — классический представитель рода Pseudomonas. Различные штаммы этих бактерий можно обнаружить всюду: в почве, воде, воздухе, гнойных ранах и сточных водах. По сравнению с другими видами псевдомонад эти формы наиболее интенсивно исследуются бактериологами, изучающими патогенную группу, и фитопатологами. Бактерии Ps. aeruginosa обладают особым разнообразием свойств, но в то же время у них отмечаются характерные общевидовые морфологические и физиологические признаки. Клетки бактерий представляют собой мелкие палочки (1,0 — 1,5 х 0,5мкм), одиночные или соединенные в пары; имеют один-два, редко три полярно расположенных жгутика, грамотрицательные. Культуры образуют синий флуоресцирующий пигмент, в состав которого входят пиоцианин. Бактерии желатин разжижают, молоко не свертывают, не пептонизируют, нитраты восстанавливают до нитритов, используют углеводы с образованием кислоты; аэробы. Температурный оптимум развития около 37 °С. Окисляют глюконат, образуя слизь. Являются сильными окислителями углеводов (сахаров, органических кислот, углеводородов). Среди этих бактерий встречаются подвижные и неподвижные формы. Многие культуры синтезируют окрашенные соединения различных типов, хорошо проникающие в субстрат. В состав пигментов входят соединения феназинового и пгеридинового ряда, а также флуоресцирующие сине-зеленые и желто-зеленые вещества.
Качественный состав пигментов и их количественный набор зависят от условий культивирования, немалое значение имеет и экологический фактор.
К этому виду близко примыкают бактерии Ps. schuilkilliensis, которые синтезируют сине-зеленый флуоресцирующий пигмент, диффундирующий в субстрат; температурный оптимум развития около 37 °С. Желатин разжижают медленно.
Культуры Ps. syncyanea образуют сине-зеленый флуоресцирующий пигмент. Желатин не разжижают, нитраты не восстанавливают; аэробы. Температурный оптимум развития около 25 °С.
Ps. scissa. Клетки мелкие (1,0x0,3 мкм), подвижные. Бактерии образуют желто-зеленый флуоресцирующий пигмент. Желатин не разжижают, молоко не изменяют, нитраты восстанавливают до нитритов; аэробы. Оптимум температуры роста около 27 °С. Эти псевдомонады были выделены из воды и почвы. К виду относятся 6 различных штаммов бактерий, обладающих сходными свойствам-и.
Ps. atlantica. Клетки мелкие, подвижные, с полярным жгутиком; колонии гладкие, блестящие. Культура энергично разжижает агар-агар и желатин, молоко пептонизирует, крахмал разлагает, нитраты не восстанавливает. Бактерии относятся к аэробам. Оптимальная температура роста около 25 °С. Эти псевдомонады были выделены из морской воды (Атлантический океан).
Ps. fluorescens. Мелкие палочки (1 —2 X 6 мкм), подвижные, имеют 2—4 полярных жгутика. Бактерии грамотрицательные. Культуры бактерий образуют зеленовато-желтый флуоресцирующий пигмент, который проникает в субстрат. Представители этого вида не синтезируют пиоцианин; хорошо развиваются на органических и синтетических средах. Колонии бесцветные или белые, выпуклые, гладкие, блестящие. Характерной особенностью этого вида является внешняя микроструктура колоний: при малом увеличении микроскопа поверхность колоний имеет характерное сетчатое или ячеистое строение. В бульоне бактерии образуют муть и пленку. Желатин разжижают, молоко не свертывают, нитраты восстанавливают до нитритов, образуют кислоту на глюкозе и сахарозе; аэробы. Оптимальная температура роста около 25 °С. Бактерии часто встречаются в воде, почве, на разных растительных и животных субстратах; не патогенны для животных.
Особое место занимает вид Ps, aurantiaса. Описание морфологических, культураль-ных и некоторых физиологических свойств бактерий этого вида встречается в основном в работах советских исследователей. Клетки бактерий этого вида палочковидные, 2—3 х 0,6 мкм, подвижные, имеют на конце 2—5 жгутиков. Колонии на питательных средах окрашены в оранжевый или красно-желтый цвет, гладкие, блестящие, плоские или выпуклые. Пигмент диффундирует в субстрат. Культуры хорошо растут на обычных питательных средах; желатин разжижают, молоко пептони-зируют. Кислоту образуют при росте на глюкозе, сахарозе, манните, глицерине. Нитраты не восстанавливают, крахмал не разлагают, аэробы. Оптимальная температура роста около 25 °С. В состав оранжевого пигмента входит несколько веществ различной химической природы.