Система И. Г. Серебрякова
Система К. Раункиера
К. Раункиер использовал для классификации жизненных форм растений единственный, но имеющий большое приспособительное значение признак – положение почек возобновления по отношению к поверхности почвы. Сначала он разработал эту систему для растений Средней Европы, но затем распространил на растения всех климатических поясов.
Все растения Раункиер разделил на пять типов (1903), в которых позднее выделил подтипы (1907).
1. Фанерофиты. Почки возобновления или верхушки побегов расположены в течение неблагоприятного времени года более или менее высоко в воздухе и подвергаются всем превратностям погоды. Подразделяются на 15 подтипов по высоте растений, по ритму развития листвы, по степени защищенности почек, по консистенции стебля. Один из подтипов – эпифитные фанерофиты.
2. Хамефиты. Почки возобновления у поверхности почвы или не выше 20–30 см. Зимой прикрыты снежным покровом. Подразделяются на 4 подтипа.
3. Гемикриптофиты. Почки возобновления или верхушки побегов на поверхности почвы, часто прикрыты подстилкой. Включают три подтипа и более мелкие подразделения.
4. Криптофиты. Почки возобновления или верхушки побегов сохраняются в почве (геофиты) или под водой (гелофиты и гидрофиты). Подразделяются на 7 подтипов.
5. Терофиты. Переносят неблагоприятное время года только в семенах.
Раункиер считал, что жизненные формы складываются исторически как результат приспособления растений к климатическим условиям. Процентное распределение видов по жизненным формам в растительных сообществах на изучаемой территории он назвал биологическим спектром. Для разных зон и стран были составлены биологические спектры, которые могли служить индикаторами климата. Так, жаркий и влажный климат тропиков был назван «климатом фанерофитов», умеренно‑холодные области имеют «климат гемикриптофитов», полярные страны – «климат хамефитов».
Критики воззрений Раункиера отмечают, что его типы жизненных форм слишком обширны и неоднородны: хамефиты включают растения с разным отношением к климату, их много как в тундрах, так и в полупустынях. И не только современный климат определяет спектр жизненных форм, но и комплекс почвенно‑литологических условий, а также история формирования флоры и влияние человеческой культуры. Тем не менее классификация жизненных форм растений по Раункиеру остается популярной и продолжает модифицироваться.
Наиболее разработанной классификацией жизненных форм покрытосеменных и хвойных на основе эколого‑морфологических признаков является система И. Г. Серебрякова (1962, 1964). Она иерархична, в ней использована совокупность большого числа признаков в соподчиненной системе и приняты следующие единицы: отделы, типы, классы, подклассы, группы, подгруппы, иногда секции и собственно жизненные формы. Собственно жизненная форма является основной единицей экологической системы растений.
Под жизненной формой как единицей экологической классификации И. Г. Серебряков понимает совокупность взрослых генеративных особей данного вида в определенных условиях произрастания, обладающих своеобразным обликом, включая надземные и подземные органы. Им выделены 4 отдела жизненных форм.
1. Отдел А. Древесные растения. Включает 3 типа: деревья, кустарники, кустарнички.
2. Отдел Б. Полудревесные растения. Включает 2 типа – полукустарники и полукустарнички.
3. Отдел В. Наземные травы. Включает 2 типа: поликарпические и монокарпические травы.
4. Отдел Г. Водные травы. Включает 2 типа: земноводные травы, плавающие и подводные травы.
Выделение отделов основано на степени одревеснения надземных осей (древесные, полудревесные и травянистые растения), выделение типов – на относительной длительности жизни надземных осей или растений в целом. Классы в пределах типов выделяются на основании структуры побегов (лиановидные, ползучие, суккулентные и проч.), на основе специфики питания (сапрофиты и паразиты) или образа жизни (эпифиты). При характеристике собственно жизненной формы растений учитывается характер надземных побегов (удлиненные, укороченные, сильно ветвящиеся и образующие подушки, ползучие и т. п.), тип корневой системы (стержнекорневые, кистекорневые, корнеотпрысковые растения и т. п.), подземные побеги (короткие и длинные корневища, клубни, луковицы, столоны, каудексы и т. п.). Учитывается также общая длительность жизни и способность к повторному цветению (монокарпики и поликарпики) и др.
Рассмотрим положение конкретных растений в системе жизненных форм И. Г. Серебрякова.
Липа сердцевидная относится к отделу древесных растений, классу кронообразующих с полностью одревесневшими удлиненными побегами, подклассу наземных, группе с подземными корнями, подгруппе прямостоячих, секции одноствольных (лесного типа), к листопадным деревьям.
Земляника лесная относится к отделу наземных трав, типу поликарпиков, классу травянистых поликарпиков с ассимилирующими побегами несуккулентного типа, подклассу столонообразующих и ползучих, группе столонообразующих, подгруппе наземностолонных. Собственную жизненную форму земляники лесной можно характеризовать как короткокорневищное кистекорневое растение с розеточными побегами и надземными столонами.
И. Г. Серебряков отмечал недоработанность и незавершенность своей классификации вследствие слабой изученности жизненных форм растений разных сообществ, особенно дождевых тропических лесов. Габитус тропических деревьев определяется зачастую не только характером стволов и крон, но и корневыми системами, поэтому последние служат важным признаком при классификации жизненных форм деревьев. Травянистые растения имеют более короткую длительность надземных осей, разнообразные ритмы сезонного развития, разный характер надземных и подземных органов. Они часто вегетативно подвижны, обладают большой семенной продуктивностью, лучше деревьев приспособлены к освоению самых разнообразных местообитаний, иногда в очень суровых условиях. Поэтому разнообразие жизненных форм у наземных травянистых растений необыкновенно велико.
Разнообразие и изменчивость жизненных форм растений. И. Г. Серебряков наметил параллельные ряды жизненных форм покрытосеменных растений и предполагаемые связи между ними (рис. 2). В сходных условиях как среди древесных, так и травянистых растений конвергентно возникли лиановидные, подушковидные, стелющиеся и суккулентные формы. Например, подушковидные формы древесные и травянистые часто встречаются в условиях хорошего освещения, но при низких температурах воздуха и почвы, при крайней сухости почвы и низкой влажности воздуха, при частых и сильных ветрах. Они обычны в высокогорьях, тундрах, пустынях, на субантарктических островах и в других местах с подобным комплексом условий.

Рис. 2. Параллельные ряды жизненных форм покрытосеменных растений и их предполагаемые связи (по И. Г. Серебрякову, 1955)
Сходные жизненные формы возникли конвергентно в разных систематических группах. Например, в условиях аридного климата пустынь одна и та же жизненная форма стеблевых суккулентов у кактусов в Америке, у молочаев и стапелий в Африке. Одну жизненную форму могут иметь как близкородственные виды (например, у манжеток), так и виды из разных семейств. К жизненной форме рыхлокустовых дерновинных поликарпиков с мочковатой корневой системой можно отнести овсяницу луговую и тимофеевку луговую (злаки), ожику волосистую (ситниковые), осоку заячью (осоковые) и т. д.
В то же время один вид может иметь разные жизненные формы. Смена жизненных форм происходит у большинства растений в онтогенезе, так как по мере роста и развития габитус меняется иногда весьма существенно. У трав стержневая корневая система часто сменяется мочковатой, розеточные побеги – полурозеточными, каудекс из одноглавого превращается в многоглавый и т. д. Иногда габитус растения закономерно меняется по сезонам. У мать‑и‑мачехи и медуницы неясной весной от корневищ отходят удлиненные, с небольшими листьями генеративные побеги. В конце мая – начале июня после плодоношения они отмирают, а из почек на корневищах у этих же особей вырастают укороченные розеточные вегетативные побеги с крупными листьями, фотосинтезирующими до осени. У безвременника великолепного каждую осень генеративное растение представлено клубнелуковицей и отходящим от нее цветком, а весной – олиственным побегом, на верхушке которого созревает плод коробочка. В подобных случаях можно говорить о пульсирующих жизненных формах.
Жизненная форма вида может меняться в пределах ареала в разных географических и экологических условиях. Многие виды деревьев на границах ареала образуют кустарниковые, нередко стелющиеся формы, например, ель обыкновенная на Крайнем Севере, ель сибирская на Южном Урале и в Хибинах.
Отдельные виды деревьев представлены разными жизненными формами в одних и тех же географических районах и даже в одних и тех же фитоценозах (рис. 3). Например, липа может быть представлена в фитоценозах: 1) одноствольным деревом; 2) порослеобразующим деревом; 3) немногоствольным деревом с 2–3 стволами; 4) многоствольным деревом – так называемое дерево‑куст; 5) куртинообразующим деревом; 6) торчками одноствольными; 7) торчками многоствольными; 8) факультативным стлаником.
В центре ареала в оптимальных условиях – на Украине, в Тульской и Пензенской областях преобладают компактные жизненные формы липы, близ северо‑восточной границы на Среднем Урале – стланиковые. Деревья‑кусты появляются после рубки одноствольных деревьев и при повреждении главной оси морозами, вредителями. Факультативный стланик входит в подлесок, приурочен обычно к сильно затененным участкам, склонам и днищам оврагов. При улучшении световой обстановки стланик может перейти в кустовидную форму или стать куртинообразующим деревом. Куртина – это заросль, образовавшаяся из одного растения. Торчки – это угнетенные низкорослые растения, выросшие при недостатке света и влаги. У молодых растений отмирают верхушки лидирующих побегов, а затем и боковых. Прожив в таком состоянии 20–30 лет, торчки могут отмереть, так и не выйдя из травяного яруса, если условия освещения улучшаются, торчки могут образовать порослевые деревья.
Другие деревья – ильм, клен, граб, черемуха и некоторые кустарники – бересклеты, жимолость, свидина, лещина и другие, также имеют широкий спектр жизненных форм. В лесах Дальнего Востока лимонник китайский в разных экологических условиях растет то как лиана, то как наземный кустарник. У травянистых растений тоже часто наблюдается внутривидовое разнообразие жизненных форм.

Рис.3. Варианты жизненной формы у липы сердцевидной (по А. А. Чистяковой, 1978):
1 – одноствольное дерево; 2 – порослеобразующее дерево; 3 – немногоствольное; 4 – многоствольное; 5 – куртинообразующее дерево; 6 – одноствольный торчок; 7 – многоствольный торчок; 8 – факультативный стланик
Экологические группы растений по отношению к воде
Гидатофиты – это водные растения, целиком или почти целиком погруженные в воду. Среди них – цветковые, которые вторично перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия, уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них редуцированы устьица и нет кутикулы. Транспирация у таких растений отсутствует, а вода выделяется через особые клетки – гидатоды.
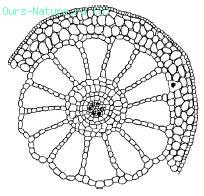
Рис. 4. Поперечный срез стебля урути Myriophyllum verticillatum (по Т. К. Горышиной, 1979)
Листовые пластинки у гидатофитов, как правило, тонкие, без дифференцировки мезофилла, часто рассеченные, что способствует более полному использованию ослабленного в воде солнечного света и усвоению СО2. Нередко выражена разнолистность – гетерофиллия; у многих видов есть плавающие листья, имеющие световую структуру. Поддерживаемые водой побеги часто не имеют механических тканей, в них хорошо развита аэренхима (рис. 4).
Корневая система цветковых гидатофитов сильно редуцирована, иногда отсутствует совсем или утратила свои основные функции (у рясок). Поглощение воды и минеральных солей происходит всей поверхностью тела. Цветоносные побеги, как правило, выносят цветки над водой (реже опыление совершается в воде), а после опыления побеги снова могут погружаться, и созревание плодов происходит под водой (валлиснерия, элодея, рдесты и др.).
Гидрофиты – это растения наземно‑водные, частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на болотах. Встречаются в районах с самыми разными климатическими условиями. К ним можно отнести тростник обыкновенный, частуху подорожниковую, вахту трехлистную, калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть эпидерма с устьицами, интенсивность транспирации очень высока, и они могут расти только при постоянном интенсивном поглощении воды.
Гигрофиты – наземные растения, живущие в условиях повышенной влажности воздуха и часто на влажных почвах. Среди них различают теневые и световые. Теневые гигрофиты – это растения нижних ярусов сырых лесов в разных климатических зонах (недотрога, цирцея альпийская, бодяк огородный, многие тропические травы и т. п.). Из‑за высокой влажности воздуха у них может быть затруднена транспирация, поэтому для улучшения водного обмена на листьях развиваются гидатоды, или водяные устьица, выделяющие капельно‑жидкую воду. Листья часто тонкие, с теневой структурой, со слабо развитой кутикулой, содержат много свободной и малосвязанной воды. Обводненность тканей достигает 80 % и более. При наступлении даже непродолжительной и несильной засухи в тканях создается отрицательный водный баланс, растения завядают и могут погибнуть.
К световым гигрофитам относятся виды открытых местообитаний, растущие на постоянно влажных почвах и во влажном воздухе (папирус, рис, сердечники, подмаренник болотный, росянка и др.). Переходные группы – мезогигрофиты и гигромезофиты.
Мезофиты могут переносить непродолжительную и не очень сильную засуху. Это растения, произрастающие при среднем увлажнении, умеренно теплом режиме и достаточно хорошей обеспеченности минеральным питанием. К мезофитам можно отнести вечнозеленые деревья верхних ярусов тропических лесов, листопадные деревья саванн, древесные породы влажных вечнозеленых субтропических лесов, летнезеленые лиственные породы лесов умеренного пояса, кустарники подлеска, травянистые растения дубравного широкотравья, растения заливных и не слишком сухих суходольных лугов, пустынные эфемеры и эфемероиды, многие сорные и большинство культурных растений. Из приведенного перечня видно, что группа мезофитов очень обширна и неоднородна. По способности регулировать свой водный обмен одни приближаются к гигрофитам (мезогигрофиты), другие – к засухоустойчивым формам (мезоксерофиты).
Ксерофиты растут в местах с недостаточным увлажнением и имеют приспособления, позволяющие добывать воду при ее недостатке, ограничивать испарение воды или запасать ее на время засухи. Ксерофиты лучше, чем все другие растения, способны регулировать водный обмен, поэтому и во время продолжительной засухи остаются в активном состоянии. Это растения пустынь, степей, жестколистных вечнозеленых лесов и кустарниковых зарослей, песчаных дюн.
Ксерофиты подразделяются на два основных типа: суккуленты и склерофиты.
Суккуленты – сочные растения с сильно развитой водозапасающей паренхимой в разных органах. Стеблевые суккуленты – кактусы, стапелии, кактусовидные молочаи; листовые суккуленты – алоэ, агавы, мезембриантемумы, молодило, очитки; корневые суккуленты – аспарагус. В пустынях Центральной Америки и Южной Африки суккуленты могут определять облик ландшафта.

Листья, а в случае их редукции стебли суккулентов имеют толстую кутикулу, часто мощный восковой налет или густое опушение. Устьица погруженные, открываются в щель, где задерживаются водяные пары.
Днем они закрыты. Это помогает суккулентам сберегать накопленную влагу, но зато ухудшает газообмен, затрудняет поступление СО2 внутрь растения. Поэтому многие суккуленты из семейств лилейных, бромелиевых, кактусовых, толстянковых ночью при открытых устьицах поглощают СО2, который только на следующий день перерабатывают в процессе фотосинтеза. Поглощенный СО2 переводится в малат. Кроме того, при дыхании ночью углеводы разлагаются не до углекислого газа, а до органических кислот, которые отводятся в клеточный сок. Днем на свету малат и другие органические кислоты расщепляются с выделением СО2, который используется в процессе фотосинтеза. Таким образом, крупные вакуоли с клеточным соком запасают не только воду, но и СО2. Так как у суккулентов ночная фиксация углекислоты и переработка ее днем в ходе фотосинтеза разделены во времени, они обеспечивают себя углеродом, не подвергаясь риску чрезмерной потери воды, но масштабы поступления углекислого газа при таком способе невелики, и растут суккуленты медленно.
Осмотическое давление клеточного сока суккулентов мало – всего 3 · 105– 8 · 105 Па (3–8 атм), они развивают небольшую сосущую силу и способны всасывать воду лишь атмосферных осадков, просочившихся в верхний слой почвы. Корневая система их неглубокая, но сильно распростертая, что особенно характерно для кактусов.
Склерофиты – это растения, наоборот, сухие на вид, часто с узкими и мелкими листьями, иногда свернутыми в трубочку. Листья могут быть также рассеченными, покрытыми волосками или восковым налетом. Хорошо развита склеренхима, поэтому растения без вредных последствий могут терять до 25 % влаги не завядая. В клетках преобладает связанная вода. Сосущая сила корней до нескольких десятков атмосфер, что позволяет успешно добывать воду из почвы. При недостатке воды резко снижают транспирацию. Склерофиты можно подразделить на две группы: эуксерофитов и стипаксерофитов.
К эуксерофитам относятся многие степные растения с розеточными и полурозеточными, сильно опушенными побегами, полукустарнички, некоторые злаки, полынь холодная, эдельвейс эдельвейсовидный и др. Наибольшую биомассу эти растения создают в период, благоприятный для вегетации, а в жару уровень обменных процессов у них очень низок.
Стипаксерофиты – это группа узколистных дерновинных злаков (ковыли, тонконоги, типчак и др.). Характеризуются низкой транспирацией в засушливый период и могут переносить особенно сильное обезвоживание тканей. Свернутые в трубочку листья имеют внутри влажную камеру. Транспирация идет через погруженные в бороздки устьица внутрь этой камеры, что снижает потери влаги.
Кроме названных экологических групп растений, выделяют еще целый ряд смешанных или промежуточных типов.
Различные пути регуляции водообмена позволили растениям заселить самые различные по экологическим условиям участки суши. Многообразие приспособлений лежит, таким образом, в основе распространения растений по поверхности земли, где дефицит влаги является одной из главных проблем экологических адаптаций.
Температурные границы существования видов
В среднем активная жизнедеятельность организмов требует довольно узкого диапазона температур, ограниченного критическими порогами замерзания воды и тепловой денатурации белков, примерно в пределах от 0 до +50 °C. Границы оптимальных температур соответственно должны быть еще более узкими. Однако реально эти границы преодолеваются в природе у многих видов за счет специфических адаптаций. Существуют экологические группы организмов, оптимум которых сдвинут в сторону низких или высоких температур.
Криофилы – виды, предпочитающие холод и специализированные к жизни в этих условиях. Свыше 80 % земной биосферы относится к постоянно холодным областям с температурой ниже +5 °C – это глубины Мирового океана, арктические и антарктические пустыни, тундры, высокогорья. Обитающие здесь виды обладают повышенной холодостойкостью. Основные механизмы этих адаптаций биохимические. Ферменты холодолюбивых организмов обладают такими особенностями строения, которые позволяют им эффективно понижать энергию активации молекул и поддерживать клеточный метаболизм при температурах, близких к 0 °C. Большую роль играют также механизмы, предотвращающие образование льда внутри клеток. При этом реализуются два основных пути – противостояние замерзанию (резистентность) и устойчивость к замерзанию (толерантность).
Биохимический путь противостояния замерзанию – накопление в клетках макромолекулярных веществ – антифризов, которые понижают точку замерзания жидкостей тела и препятствуют образованию кристаллов льда в организме. Такого типа холодовые адаптации обнаружены, например, у антарктических рыб семейства нототениевых, которые живут при температуре тела ‑1,86 °C, плавая под поверхностью сплошного льда в воде с такою же температурой. Мелкая тресковая рыба сайка в Северном Ледовитом океане плавает в водах с температурой не выше +5 °C, а нерестится зимой в переохлажденных водах у побережья. Глубоководные рыбы в приполярных районах все время находятся в переохлажденном состоянии.
Предельная температура, при которой еще возможна активность клеток, зафиксирована у микроорганизмов. В холодильных камерах мясные продукты могут быть испорчены за счет деятельности бактерий при температурах до ‑10‑12 °C. Ниже этих температур роста и развития одноклеточных организмов не происходит.
Другой путь холодостойкости – выносливость к замерзанию – связан с временным прекращением активного состояния (гипобиозом или криптобиозом).Образование кристалликов льда внутри клеток необратимо нарушает их ультраструктуру и приводит к гибели. Но многие криофилы способны переносить образование льда во внеклеточных жидкостях. Этот процесс приводит к частичной дегидратации клеток, что повышает их устойчивость. У насекомых накопление защитных органических веществ, таких как глицерин, сорбит, маннит и других, препятствует кристаллизации внутриклеточных растворов и позволяет переживать критические морозные периоды в состоянии оцепенения. Так, жуки‑жужелицы в тундрах выдерживают переохлаждение до ‑35 °C, накапливая к зиме до 25 % глицерина и снижая содержание воды в теле с 65 до 54 %. Летом глицерин в их теле не обнаруживается. Некоторые насекомые выдерживают зимой до ‑47 и даже ‑50 °C с замерзанием внеклеточной, но не внутриклеточной влаги. Морские обитатели практически не сталкиваются с температурами ниже ‑2 °C, но беспозвоночные приливно‑отливной зоны (моллюски, усоногие раки и др.) зимой во время отлива переносят замерзание до – (15–20) °С. Клетки под микроскопом выглядят сморщенными, но кристаллов льда в них не обнаруживается. Устойчивость к замерзанию может проявляться и у эвритермных видов, оптимальные температуры развития которых далеки от 0 °C.
Термофилы – это экологическая группа видов, оптимум жизнедеятельности которых приурочен к области высоких температур. Термофилией отличаются многие представители микроорганизмов, растений и животных, встречающихся в горячих источниках, на поверхности прогреваемых почв, в разлагающихся органических остатках при их саморазогревании и т. п.
Верхние температурные пределы активной жизни отличаются у разных групп организмов. Наиболее устойчивы бактерии. У одного из видов архебактерий, распространенных на глубинах вокруг термальных источников («курильщиков»), экспериментально обнаружена способность к росту и делению клеток при температурах, превышающих +110 °C. Некоторые бактерии, окисляющие серу, как, например, Sulfolobus acidocaldarius, размножаются при +(85–90)°С. Обнаружена даже способность ряда видов расти в практически кипящей воде. Естественно, не все бактерии активны при столь высоких температурах, но разнообразие таких видов достаточно велико.
Верхние температурные пороги развития цианобактерий (сине‑зеленых водорослей) и других фотосинтезирующих прокариот лежат в более низких пределах от +70 до +73 °C. Термофилы, растущие при +(60–75) °С, есть как среди аэробных, так и анаэробных бактерий, спорообразующих, молочнокислых, актиномицетов, метанообразующих и др. В неактивном состоянии спорообразующие бактерии выдерживают до +200 °C в течение десятков минут, что демонстрирует режим стерилизации предметов в автоклавах.
Термостабильность белков бактерий создается за счет значительного числа малых изменений в их первичной структуре и добавочных слабых связей, определяющих укладку молекул. В транспортных и рибосомных РНК термофилов повышено содержание гуанина и цитозина. Эта пара оснований более термостабильна, чем пара аденин – урацил.
Таким образом, выход температурной устойчивости за пределы средней нормы происходит в основном за счет биохимических адаптаций.
Среди эукариотных организмов – грибов, простейших, растений и животных – также существуют термофилы, но уровень их толерантности к высокой температуре ниже, чем у бактерий. Пределы роста грибного мицелия составляют +(60–62) °С. Известны десятки видов, способных быть активными при +50 °C и выше в таких местообитаниях, как компосты, стога сена, хранящееся зерно, прогреваемая почва, свалки и т. п. Простейшие – амебы и инфузории, одноклеточные водоросли могут размножаться до температуры в +(54–56) °С Высшие растения могут переносить краткосрочные нагревания до +(50–60) °С, но активный фотосинтез даже у пустынных видов тормозится температурами, превышающими +40 °C. Так, в клетках суданской травы при +48 °C движение цитоплазмы останавливается уже через 5 мин. Критические температуры тела некоторых животных, например пустынных ящериц, могут достигать +(48–49) °С, но для большинства видов температуры тела, превышающие +(43–44) °С, несовместимы с жизнью из‑за рассогласования физиологических процессов и коагуляции белка коллагена. Таким образом, с усложнением организации живых существ способность их быть активными при высоких температурах понижается.
Узкая специализация и латентные состояния намного раздвигают границы жизни по отношению к отдельным факторам среды. Если средние температурные пределы активности организмов характеризуются диапазоном от 0 до +(40–45) °С, то специализированные виды (криофилы и термофилы) расширяют его более чем вдвое (от ‑10 до примерно +110 °C), а в состоянии криптобиоза и анабиоза некоторые формы жизни способны выдерживать температуры, близкие к абсолютному нулю или намного превышающие точку кипения виды.