На примере аборигенов Австралии и Океании 17 страница
Различия в уровне таксономической обособленности современных голоэпипелагических рыб и прибрежных родственных им форм показывают, что колонизация океанской эпипелагиали представляет собой перманентный, очень растянутый во времени процесс, продолжающийся, судя по встречаемости в открытом океане видов, общих с неритической зоной, до настоящего времени. В этом случае мезоэпипелагические и ксеноэпипеелагические рыбы должны рассматриваться в качестве очередных «разведывательных групп», ведущих освоение открытого океана.
Трудно, пожалуй, вообразить себе рыбу, менее соответствующую классическому представлению об обитателе эпипелагиали, чем диодоны, или рыбы-ежи, из рода двузубов. Все они очень похожи друг па друга. Их короткое, на первый взгляд довольно неуклюжее тело покрыто длинными острыми колючками, которые плотно прижимаются к коже при спокойном плавании. Диодоны способны раздуваться до шарообразной формы, наполняя воздухом или водой особый мешок (вырост желудка), а их колючки при этом встают торчком, как у настоящих ежей, и превращаются в эффективное оружие пассивной защиты. Двузубы - типичные обитатели прибрежных мелководий; их зубы преобразованы в сплошные твердые пластины (по одной в каждой челюсти — отсюда и название этих рыб), очень полезные для разгрызания иглокожих, моллюсков, кораллов. В то же время мальки некоторых рыб-ежей довольно обычны в эпипелагиали, а один вид - океанический двузуб (Diodon eydouxii), внешне мало отличающийся от других сородичей, постоянно живет в открытом океане, и мощные зубные пластины нисколько не мешают ему питаться мелким планктоном. Диодонов часто удается наблюдать на световых станциях, и, должен сказать, в воде эти рыбы отнюдь не выглядят очень уж неуклюжими. Они с большим проворством пересекают освещенную зону и довольно ловко уклоняются от закидного сачка, но даже легкое прикосновение к ежу вызывает у него совершенно неадекватную реакцию: он немедленно превращается в колючий шар, который, может быть, трудно проглотить хищнику, но очень просто достать из воды любой сеткой.
На мой взгляд, океанический двузуб самим фактом своего существования представляет одно из наиболее наглядных доказательств продолжающегося заселения эпипелагиали прибрежными рыбами. Это — пример недавнего вселенца, нашедшего в новом биотопе благоприятные условия, но еще не успевшего приобрести специфических адаптации к нему. На противоположном полюсе я поместил бы луваря (Luvarus imperialis) (рис. 43) — удивительную, внешне ни на кого не похожую рыбу, которую в продолжение многих лет считали родственной скумбриевым или ставридовым. Новейшие анатомические исследования показали, что луварь ведет начало от прибрежных рыб-хирургов, но длительная эволюция в условиях открытого океана сделала его совсем непохожим па своих предков.
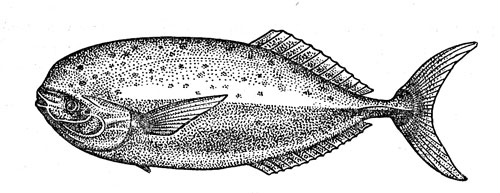
Рис. 43. Луварь - рыба, родственные связи которой долго оставались непонятными.
В эпипелагиали явно преобладают филогенетически молодые группы костистых рыб (в частности, окунеобразные). Основываясь на этом факте, я предполагал ранее, что до появления этих вселенцев верхние слои открытого океана были заселены низшими (мягкоперыми) рыбами, которые, не выдержав конкуренции, оказались вытесненными вниз и дали начало современному населению средне-глубинной зоны. Сейчас я считаю это умозрительное предположение крайне маловероятным. Существует гораздо больше оснований для иной точки зрения: главным источником формирования мезопелагического ихтиоцена послужили придонные и придонно-пелагические формы, обитавшие на материковом склоне.
Представление о происхождении мезопелагических рыб от мезобентальных и мезобентопелагических предков основывается на следующих соображениях. Специфические подотряды и семейства мезопелагических рыб относятся к таким отрядам (угреобразные, аулопообразные, миктофообразные, трескообразные, бериксообразпые и др.), основная масса представителей которых обитает на континентальном склоне, и являются наиболее специализированными таксонами этих групп. Семейство Chiasmodontidae наиболее родственно верхнемезобенталыюму сем. Champsodontidae. Мезопелагические роды в семействах, распространенных и вне рассматриваемого биотопа, принадлежат к группам, характерным в основном для мезобеитали (Isistius и Euprotomicrus — к сем. колючих акул, Хеnорhthalmichthys — к сем. серебрянковых, Scopelengys — к сем. неоскопеловых, Brotulotaenia — к сем. ошибневых, Супо-macrurus и Mesobius — к сем. долгохвостовых, Diretmus — к сем. диретмовых, Diplospinus и Nealotus — к сем. гемпиловых, Ectreposebastes — к сем. скорпеновых, Nectoliparis и Rhodichthys — к сем. морских слизней). В то же время мезопелагические виды в преимущественно придонных родах, по-видимому, очень редки: единственный известный мне пример представляет Nezumia parini, — описанный К. Хаббсом и Т. Ивамото пелагический долгохвост из восточной тропической Пацифики.
Бати- и абиссопелагический ихтиоцены в основном формировались, по-видимому, тем же путем, что и мезопелагический. Судя по распространению рецентных родственных групп, специфические высокоспециализированные глубоководные семейства угреобразных (пилосошниковые, циемовые, мешкоротовые) и удильщикообразных (11 семейств подотряда Ceratioidei) ведут свое начало от донно-придонных рыб нижней части материкового склона. В пелагиали больших глубин значительная роль принадлежит и представителям таких семейств, которые еще не полностью утратили связь с дном — гладкоголовам и платитроктовым, откладывающим донную икру, а также долгохвостам, афионидам и др. В то же время наличие в составе глубинной пелагической ихтиофауны родов, принадлежащих к преимущественно мезопелагическим семействам гоностомовых, малакостеевых и миктофовых, указывает на частичное заселение этой зоны из средних горизонтов.
Нужно сказать, что представители отдельных групп мезопелагических рыб, принадлежащих к макропланктону, оказались способными выйти за пределы исходного биотона и значительно расширить адаптивную зону обитания этих групп. Наиболее характерные примеры внедрения во все смежные участки среды дают светящиеся анчоусы (сем. миктофовые) которые справедливо считаются одной из самых процветающих групп мезопелагического ихтиоцена. Тем не менее около трети видов семейства (а их число достигает 220) временно или постоянно входит в состав других сообществ. Так, около 40 видов поднимаются ночью в эпипелагиаль (по меньшей мере один из них — Benthosema pterotum, наблюдавшийся в восточной тропической Пацифике у поверхности даже в светлое время суток, может образовывать там жилую, т. е. не мигрирующую, популяцию), около 10 видов живут в батипелагиали и около 20 — вторично возвратились в бентопелагические биотопы (туда, где когда-то обитали предковые формы миктофид) и в некоторых случаях достигли там высокой численности. Вторичнобентопелагические виды возникли и в других мезопелагических семействах — среди топориковых (достаточно вспомнить мавролика), фотихтовых, гоностомовых, меланостомиевых, меламфаевых. Такие виды есть также среди мезобатипелагических, в основной своей массе глубоководных удильщиков церациоидей: это представители родов Diceratias и Phrynichthys. Мало того, один из видов сем. тауматихтовых — Thaumatichthys axeli является, по всей вероятности, вторичнодонной рыбой (единственный в своем роде случай «бентизации», контрастирующий с обычной «пелагизацией» генеративно донных форм).
Говоря о заселении рыбами океанических биотопов, нельзя обойти вниманием вопрос о параллелизмах и конвергенциях в их эволюции, сопровождавших освоение тех или иных участков среды обитания. Этот вопрос, издавна привлекавший внимание исследователей, был в недавнее время детально разобран Н. Маршаллом, что позволяет ограничиться здесь рассмотрением отдельных, наиболее ярких проявлений функционального и структурного сходства систем и органов у рыб, входящих в состав разных сообществ. В эпипелагиали наиболее поражает разительное сходство между акулами ламнидами (к ним относятся, напомню, сельдевые акулы, мако и кархародон) и тунцами. Это касается не только общей формы идеально обтекаемого торпедообразного тела, его окраски, положения и относительных размеров непарных плавников, наличия стабилизирующих килей на чрезвычайно узком хвостовом стебле, несущем мощный хвостовой плавник в форме полумесяца. Еще более замечательны конвергенции в строении внутренних органов, и главная из них несомненно состоит в необычайном развитии подкожной сосудистой системы, питающей туловищную мускулатуру и обеспечивающей «теплокровность» у этих совсем не родственных рыб. И у тунцов и у ламповых акул мощно развиты наиболее активные красные мышцы латеральной мускульной системы. Волокна этих мышц окружены «чудесной сетью» капилляров, которая обеспечивает интенсивный теплообмен между артериальной кровью, исходно (в жабрах) имеющей температуру морской воды, и венозной кровью, нагретой за счет мышечной работы. В результате этого обмена температура тела тунцов и акул оказывается па 6—10° выше «забортной» температуры, и это приводит к значительным энергетическим выгодам (ведь скорость всех ферментативных реакций повышается в таком случае в 2—3 раза). Крупные тунцы (виды рода Thunnus) и акулы сем. Lamnidae могут считаться «идеальными нектерами» — как по «крейсерским» достоинствам, так и по максимальной скорости на спринтерской дистанции они не имеют соперников ни среди других рыб (исключение составляют мечевидные, достигшие тех же результатов), ни среди кальмаров и морских млекопитающих.
Примеры конвергенции и параллелизмов среди глубоководных пелагических рыб очень многочисленны, и их полное перечисление заняло бы здесь слишком много места. Нельзя не упомянуть тем не менее бросающегося в глаза сходства в окраске тела — серебристого у многих верхыемезопелагических видов (из батилаговых, топориковых, миктофовых, палочкохвостых и др.) и черного или коричневого у всех форм, обитающих на глубине более 1000 м. Органы свечения независимо возникли у колючих акул (отряд катраыообразные) и у костистых рыб, принадлежащих к отрядам лососеобразных, стомиеобразных, аулопообразных, миктофообразных, окунеобразных (род Pseudoscopelus). Телескопические глаза характерны для представителей отрядов лососеобразных (Xenophthalmichthys, Opisthoproctus), аулопообразных (Gigantura, Scopelarchus, Evermannella), миктофообразных (Protomyctophum), опахообразных (Stylephorus). Многие стомиеобразные и все удильщики приманивают добычу своеобразными удочками со светящейся приманкой. Для всех хищников характерны приспособления для захвата крупной добычи — огромные пасти, зубатые челюсти, редуцированные жаберные тычинки, сильно растяжимые желудки и стенки тела. У идиакантов и удильщиков карликовые самцы значительно мельче самок. Параллельным путем возникли и вертикальные миграции мезопелагических рыб.
Многие рыбы, обитающие в придонных слоях (галозавры, долгохвосты, ателеопы, ошибневые), независимо приобрели очень сходный тип строения, для которого характерны выступающее рыло, нижний рот, очень длинная, постепенно сходящая на нет хвостовая часть туловища, несущая по нижнему краю многолучевой аномальный плавник. Ундуляции такого плавника переводят рыбу в наклонное (головой вниз) положение, очень удобное для поиска корма на грунте. Все эти рыбы обладают плавучестью, близкой к нейтральной, как, впрочем, и ведущие сходный образ жизни придонные колючие акулы, но конвергентно сходный результат достигается принципиально разными путями — все перечисленные костистые рыбы имеют плавательный пузырь, а хрящевые регулируют плавучесть высоким содержанием легкого жира в огромной печени.
Скудность палеонтологического материала, значительные в ряде случаев отличия океанических таксонов рыб от родственных прибрежных форм, а также их сходство между собой, определяемое конвергенциями и параллелизмами, создают определенные трудности для воссоздания эволюционной истории рыб открытого океана. Тем не менее происхождение рецептной океанической ихтиофауны вполне очевидно: исходным материалом для ее формирования были шельфовые и неритические предковые формы. Заселение придонных глубин и открытой пелагиали, осуществлявшееся многими группами независимо друг от друга, представляется очень растянутым во времени. Датировка начала этого процесса, теряющаяся в далеком прошлом, невозможна без малооправданных спекуляций, а на примере эпипелагиали и мезобентали видно, что заселение основных океанских биотопов продолжается и поныне. В результате для каждого из этих биотопов характерна разновозрастная фауна.
Как известно, степень морфо-экологической специализации к условиям глубин, косвенно отражающая филогенетический возраст таксонов, была положена А. П. Андрияшевым в основу разделения глубоководных рыб на две группировки — древнеглубоководную и вторичноглубоко-водную. Такое разделение, несомненно реальное и очень наглядное в некоторых крайних случаях, все же довольно относительно и не дает возможности альтернативно определить принадлежность любой и каждой систематической группы рыб к одной из этих категорий. Представляется, что наиболее резко обособлены от ближайших родственников таксоны, возникшие в результате пелагизации, т. е. при переходе от бентического образа жизни к бентопелагическому, а затем к обитанию в безбрежной толще вод. Эти переходы носившие прогрессивный характер, сопровождались повышением общего уровня организации и соответствовали, таким образом, введенному академиком А. Н. Северцовым понятию «ароморфоз», тогда как постепенные адаптивные преобразования при увеличении глубины обитания не выходили за рамки частных приспособлений.
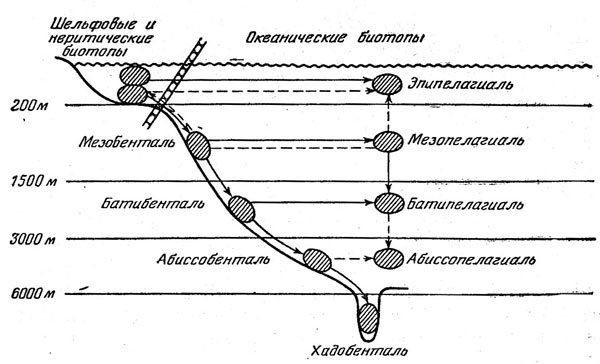
Рис. 44. Пути заселения океанических биотопов рыбами.
Генеральная схема заселения океанических биотопов представляется сейчас в следующем виде (рис. 44). Шельфовые предки дали начало мезобентальным рыбам, а от них последовательно произошли бати- и абиссобентальные и хадальные формы. В этом процессе очень существенная роль принадлежала вытеснению на большие глубины примитивных представителей тех или иных таксонов, не выдержавших конкуренции с более прогрессивными формами. Заселение разных горизонтов пелагиали происходило от придонных и неритических предков независимо на разных глубинах и шло главным образом по пути прогрессивной ароморфной эволюции; в формировании батипелагической фауны существенное значение имело и заселение сверху, из мезопелагиали. Картина современного биотопического распределения океанических рыб осложняется вторичным подъемом представителей некоторых групп в более поверхностные горизонты, вторичным же возвращением из открытой пелагиали к склонам.
В начале этой книги я упомянул, что к числу объектов промысла относятся сотни видов рыб. «Тысячи,— поправил Н. П. Новиков, мой давний приятель, а ныне директор АзЧерНИРО - рыбохозяйственного института в Керчи,— ведь в "Словаре названий морских промысловых рыб" Г. У. Линдберга и соавторов значится три с лишним тысячи таких видов». Я задумался. Что же такое «промысловые рыбы» и не слишком ли расширенно мы толкуем это понятие? В упомянутом «Словаре», как сказано в предисловии к нему, «приводятся по возможности все виды рыб, служащие объектами рыболовства, а также обычные в уловах и имеющие народные названия». Это определение не отличается строгостью и к тому же представляется неправильным по своему существу - ведь многие рыболовные орудия облавливают водоемы безвыборочно, а народные названия есть почти у всех рыб, распространенных в пределах территории, которую этот народ занимает. Кстати говоря, несъедобных рыб практически не существует (кого только не приходилось дегустировать в экспедициях!), хотя некоторые из них могут употребляться в пищу только приготовленными соответственным образом. Я думаю, что в наше время «промысловыми рыбами» следует называть только тех, которые образуют локальные концентрации, достаточные для экономически оправданного облова, а отнесение к их числу рыб, случайно попадающих в снасти (так называемого «прилова»), неправомочно. К «потенциально промысловым рыбам» принадлежат такие виды, которые имеют достаточно большую биомассу, но не могут рентабельно облавливаться при современном состоянии промышленного рыболовства.
Промысловые и потенциально промысловые рыбы отличаются от не промысловых прежде всего высокой численностью и большей плотностью популяций. Отсюда следует, что они относятся к числу доминирующих видов своих ихтиоценов. Исключение составляют крупные хищники, занимающие в трофических пирамидах самые верхние уровни: их промысловое значение определяется не количеством, а качеством — очень высокой ценностью получаемого сырья. Доминирующие виды считаются оптимально приспособленными к конкретным условиям сообществ, хотя в чем именно проявляется эта приспособленность — сказать трудно. К примеру, из нерито-пелагических рыб (почти все они выходят и в открытые воды) преобладающее большинство — близкие подвиды или виды из родов Clupea (сельди), Sardinops (сардины иваси), Engraulis (анчоусы), Micromesistius (путассу), Trachurus (ставриды), Scomber (скумбрии) — имеют промысловое значение во всех районах, где они встречаются. Это правило не распространяется, однако, на род Theragra (минтай). Тихоокеанский минтай (Т. chalcogramma) принадлежит к числу важнейших промысловых рыб (в последние годы он твердо удерживает первое место в мире по вылову), а почти неотличимый от него атлантический вид (Т. finmarchica) остается известным по 4 коллекционным экземплярам, пойманным в юго-западной части Баренцева моря.(Т. С. Расе предполагает, что атлантического вида минтая не существует, а относимые к нему особи представляют собой тихоокеанских рыб, достигших берегов Норвегии «северным морским путем» в теплый год.)
Известно также, что виды рыб могут с легкостью переходить из разряда непромысловых в категорию промысловых и обратно. Такие переходы могут быть следствием естественных флюктуации численности («волн жизни»), антропогенного воздействия, в том числе перелова, или природных катаклизмов. Хорошо известным примером такого катаклизма служит почти полное вымирание в 1882 г. хамелеоноголового хохлача (Lopholatilus chamaeleoniceps) (сем. малакантовые), обитающего у атлантического побережья США в водах верхней части континентального склона (глубина 100—300 м). Хохлач живет в очень узком диапазоне температуры (8—12°), так что даже относительно небольшое охлаждение воды привело тогда к его массовой гибели (по приближенной оценке на поверхности было насчитано около 1,5 млрд мертвых рыб) и вид надолго выбыл из списка промысловых объектов. Случаи массовой смерти придонных рыб наблюдались и в других районах океана.
Современное океаническое рыболовство (т. е. рыболовство за пределами шельфов) имеют недолгую историю. В крупных масштабах оно стало развиваться только в последние годы, но есть все основания считать, что локальный промысел океанических объектов в некоторых районах существовал издавна. Это касается в первую очередь тех участков, где склон начинается в непосредственной близости от берега. В Полинезии, например, существовали (а на некоторых островах существуют и сейчас) традиционные способы лова многих рыб открытого океана — летучек, тунцов, корифен, руветты, акул. Для каждого объекта были разработаны специальные приемы и методы промысла, основанные на прекрасном знании повадок этих рыб. Летучих рыб с незапамятных времен ловили там, используя их положительную реакцию на искусственный свет: в темные ночи их привлекали к лодкам горящими факелами. При ужении мелких тунцов островитяне применяли снасти, изготовленные из перламутровых раковин жемчужниц. Такая «блесна» имеет форму слегка изогнутой стилизованной рыбки длиной 8—10 см, к хвосту которой привязывается под небольшим углом костяное, деревянное или каменное острие, заменяющее крючок, а сзади прикрепляется пучок перьев или свиной щетины. Обнаружив стаю мелких тунцов или пеламид (рыб ищут, наблюдая за их пищевыми конкурентами — морскими птицами) и приблизившись к ней, рыбак тянет блесну за лодкой, а при поклевке тупца резким рывком, продолжающим его движение в воде, выбрасывает добычу в каноэ. Нужно добавить, что полинезийцам прекрасно известна привычка мелких рыбешек собираться у плавучих предметов, где их и находят тунцы. Поэтому, увидев плавник — бревно, ветки или что-нибудь подобное, рыбаки забирают его в свое каноэ, под которое переходят укрывавшиеся там рыбки, после чего спокойно ожидают появления тунцов или корифен. Крупных тунцов и марлинов ловят на наживленные крючки, иногда с использованием живой приманки, сохраняющейся в плавучих корзинах. Особенно специфичен промысел руветты (местное название «уравена», или «куравена»), при котором попадаются и другие хищные рыбы, обитающие в верхней части склона. Для этого рыбаки выходят в океан ночью и опускают на дно на глубину около 200 м наживку, насаженную на своеобразный деревянный крючок довольно сложной конструкции (эти крючки, как и тунцовые «блесны», имеют свою специфику на разных островах Океании).
Кустарные и полукустарные способы лова океанических рыб до сих пор практикуются во многих районах тропической зоны, а кое-где даже сохраняют в слегка измененном виде свое былое значение. На юге Вьетнама, например, важную роль играет традиционный промысел летучих рыб. В апреле 1981 г. я был в этой стране и проехал на машине около 600 км вдоль побережья Южно-Китайского моря от Нячанга (там находится Институт морских исследований ДРВ) до Дананга. На этом пути мы посетили несколько рыбацких кооперативов и побывали на множестве рынков, расположенных у причалов на самом берегу. Туда, непосредственно к прилавкам, в ассортименте подвозят по утрам свой улов рыбаки-единоличники. «Инспекция» рынков меня прямо-таки поразила: везде и всюду доминировали летучие рыбы (удалось собрать отличную коллекцию — 12 разных видов), которых продавали и в свежем и в вяленом для хранения впрок виде. Вялят их, кстати, очень простым способом, рассыпая без всякой предварительной обработки под жарким солнцем на асфальтированных или хорошо утрамбованных земляных площадках. Думаю, летучки идут и на изготовление своеобразного соуса «нуок-мам», получаемого длительным (в течение трех-четырех месяцев) квашением пересыпанной солью рыбы,- вьетнамцы употребляют его как белковую приправу к рису и супу. Рыбу подвозят к рынкам на моторных лодках, которые, мне сказали, уходят для промысла на 10—20 км от берега. На лове летучек вьетнамские рыбаки используют очень низкие (всего 1,5—2 м) плавные сети, выставляемые на ночь длинными порядками.
Согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), содержащей официальные данные об уловах всех государств мира, океанические рыбы добываются многими странами. В перечне промысловых объектов значатся, в частности, ламповые, колючие и серые акулы, ромбовые скаты, лососи, серебрянки, мавролик, светящиеся анчоусы, зеленоглазки, макрелещука и сайра, летучие рыбы, моровые, хэки (мерлузы), долгохвосты, рыба-бекас, берикс, большеголовы, солнечники, ставридовые, красноглазки, белокровные рыбы, рексии, тунцы, скумбрии, марлины, копьеносцы, меч-рыба, беспузырные окуни, угольная рыба, палтусы и многие другие обитатели открытого океана. По данным «Ежегодников ФАО», мировой вылов в 1978—1984 гг. составлял 70,1—82,8 млн т (из которых 63,1— 73,1 млн т добывалось в морях и океанах) и постоянно увеличивался из года в год. В этот же период улов СССР, прочно удерживающего первое место в мире по вылову рыбы, вырос с 9,0 до 10,6 млн т (в морских водах с 8,3 до 9,7 млн т). При этом доля океанических объектов в мировом улове не достигает и 10%. В советском рыболовстве она превышает 14%, но подавляющая часть этого улова (10—11%) приходится на псевдонеритических рыб, добываемых как в открытых водах, так и в неритической зоне.
Рассматривая уловы отдельных объектов океанического рыбного промысла, нужно прежде всего отметить, что имеющаяся статистика не дает возможности различать в уловах псевдонеритических рыб части, взятые в неритических и открытых водах, и это заставляет отказаться от попыток анализа соответствующих цифр. Среди голоэпипелагических рыб наибольшие уловы обеспечивали в 1978—1984 гг. тунцы рода Thunnus, в основном желтоперый, большеглазый и длинноперый (1017—1059 тыс. т), полосатый тунец (712—1050 тыс. т), сайра (200—460 тыс. т), мелкие тунцы (153—212 тыс. т), летучие рыбы (44— 74 тыс. т), копьеносцы и марлины (57—63 тыс. т), меч-рыба (39—63 тыс. т), корифены (18—29 тыс. т), морские лещи (4—10 тыс. т) и сельдевые акулы (1—4 тыс. т). Правда, уловы прочих пелагических акул, включенных в рубрику «Carcharhinidae», составляет 35—47 тыс. т, и львиная доля этого вылова приходится, конечно, на синюю и длиннокрылую акул. В число важнейших объектов промысла на материковом склоне входят хэки (мерлузы), которых вылавливали от 1117 до 1675 тыс. т (из 11 промысловых видов рода Merluccius наибольшие уловы дают капская, аргентинская, орегонская и восточноатлантиче-ская мерлузы), морские окуни (322—489 тыс. т), палтусы (130—161 тыс. т), морские щуки (род Molva; 80—93 тыс. т), морские черти (48—89 тыс. т), долгохвосты (17— 67 тыс. т), моровые рыбы (в основном лемонема) (2— 59 тыс. т), аргентины (17—36 тыс. т), угольная рыба (20— 33 тыс. т), рыба-бекас (9—32 тыс. т) и глубоководные солнечники, или лунники (6—28 тыс. т). Нужно иметь в виду, впрочем, что часть этих уловов несомненно получена на глубине менее 200 м.
Каждая океаническая рыба хорошо «знает свое место», и в естественной среде многие из них никогда не могут встретиться друг с другом. Возможность ненадолго оказаться в одной точке пространства сохраняют, однако, промысловые рыбы, но для этого они должны сначала попасть в руки японских рыбаков. Дело в том, что значительная часть их уловов, полученных в разных районах Мирового океана, поступает потом на оптовый рыбный рынок Токио - гигантское торговое предприятие, через которое в 60-е годы проходило 15—20% общего улова страны (напомню, что Япония занимает первое место в мире по валовому вылову морепродуктов и ненамного уступает СССР по вылову собственно рыбы).
...Когда «Витязь» посетил Токио в 1964 г., нас взял под опеку профессор Токихару Абэ, хорошо известный не только своими многочисленными научными публикациями, но и неизменной доброжелательностью по отношению к коллегам-ихтиологам из любой страны. Будучи сотрудником университета, профессор Абэ в то время занимал также пост инспектора Токийского оптового рынка: ежедневно в 5 часов утра он должен был просматривать подготовленные для аукциона лоты (партии «товара»), чтобы не допустить продажу ядовитых или подозрительных в этом отношении рыб. Я с радостью согласился посетить рынок вместе с ним, и впечатление от увиденного до сих пор остается очень ярким. Рыбы, доставленные из открытого океана, конечно, составляли небольшую часть «экспозиции», но были представлены во всем своем многообразии. Огромные туши марлинов и меч-рыб лежали рядами, соседствуя с целыми «поленницами» крупных тунцов и акул. Штабелями высились аккуратные ящики, коробки, корзины с лососями, мелким тунцом, скумбрией, сардиной, сайрой, летучими рыбами. В некоторых сборных партиях «морской рыбы» можно было увидеть и довольно редких представителей океанической фауны (помню, например, алепизавров, опаха, каких-то долгохвостов и морид). Недаром многие коллекционные экземпляры п музеях многих стран мира (в том числе и в Зоологическом музее АН СССР в Ленинграде) имеют одинаковые этикетки («рыбный рынок в Токио»), различающиеся только датой поступления.
В соответствии с разнообразием промысловых объектов в открытом океане набор методов океанического рыболовства очень широк. Здесь находят применение траловый, кошельковый, ярусный, удебный, электросветовой, дрифтерный способы лова, применение которых определяется специфическими особенностями экологии и поведения промысловых рыб.
Важнейшее значение в современном промысле принадлежит, пожалуй, траловому рыболовству, дающему возможность активного и очень производительного облова скоплений рыб на дне, у дна и в толще воды в значительном удалении от грунта. Сейчас промыслом используются различные модификации так называемых оттертралов, которые раскрываются в воде за счет гидродинамических сил, возникающих при их буксировке. Этот эффект достигается применением распорных досок, которые прикрепляются к крыльям трала под некоторым углом к направлению его движения. Габариты тралов ограничиваются только размерами судна (траулера) и мощностью его главного двигателя и могут быть очень большими. Новейшие модели канатных разноглубинных тралов имеют, например, вертикальное и горизонтальное раскрытие порядка 70 м. При скорости хода с тралом, достигающей 6 узлов (10,8 км/ч), они облавливают, таким образом, до 12 тыс. м3/с, и улов в 30—40 т представляется вполне, обычным. Лов разноглубинным тралом производится прицельно, т. е. по показаниям гидроакустических приборов, регистрирующих косяки рыбы. Применяемая контрольная аппаратура позволяет постоянно следить также за раскрытием трала, горизонтом его хода, накоплением улова в траловом мешке. В океаническом рыболовстве применяют в основном разноглубинные тралы, используемые в пелагическом или придонном варианте (работы с донными тралами возможны в немногих районах, например на вершинах гайотов). Основными объектами тралового промысла служат стайные псевдонеритические рыбы — сардина иваси, сельдь, скумбрия, ставрида и др., а также рыбы, приуроченные в своем распространении к материковому склону (лемонема, долгохвосты, палтусы, угольная рыба) и подводным поднятиям (бе-рикс, мавролик, рыба-кабан, лунник).
Кошельковый промысел ведется со специальных скоростных и маневренных судов — сейнеров. Применяемое орудие лова (кошельковый невод) представляет собой длинную (до 1,5—1,8 км) и высокую (до 200—300 м) сеть, верхняя подбора которой снабжена поплавками, а нижняя — грузилами и большими металлическими кольцами (через них продевается стягивающий трос). Обнаружив у поверхности косяк рыбы (для этого нередко используют палубный вертолет), сейнер спускает на воду моторную шлюпку и обметывает стаю неводом, полностью замыкая круг, затем стягивающий трос быстро берется на лебедку и нижняя кромка невода затягивается «кошельком», не давая рыбе уйти вглубь. Современные океанские сейнеры используются главным образом для лова тунцов, скумбрии и других подвижных рыб с высокой скоростью плавания.