На примере аборигенов Австралии и Океании 9 страница
Функция системы светящихся органов, по-видимому, многозначна, но главное ее назначение состоит, без сомнения, в обеспечении распознания особей своего вида, а в некоторых случаях и их пола. Сосредоточение светопроду-цирующих источников преимущественно на нижней части тела позволяет предполагать, что здесь, имеет место и так называемый противотеневой эффект, способствующий лучшей маскировке рыбы на фоне рассеянного верхнего освещения. Есть также предположения, что вспышки хвостовых желез могут дезориентировать преследующего хищника.
Систематика миктофид, которую нередко вполне серьезно именуют «миктофидологией»,- очень своеобразный раздел ихтиологической науки. Миктофидологи (в нашей стране эта специфическая профессия полноправно представлена В. Э. Веккером) определяют роды и виды, рассматривают их родственные отношения, строят систему семейств, основываясь главным образом на расположении фотофоров, каждую группу которых принято обозначать определенной буквенной аббревиатурой, а отдельные органы в ее составе — порядковыми номерами. Поэтому научные рисунки светящихся анчоусов часто напоминают технические чертежи, а видовые описания типа «SAOi под V04 или впереди его вертикали; SA02 примерно на вертикали AOa1; SAO3 над АОа2 и т. д. понятны «без перевода» только тем, кто хорошо знаком с этой группой рыб.
Такая «геометричность» миктофид, казалось бы, очень облегчает их опознание: на схематических рисунках различия между видами очень наглядны и очевидны. Это, однако, вовсе не так. На самом деле в уловах глубоководных тралов крайне редко можно встретить идеально сохранившийся экземпляр с полным набором светящихся органов,- а установление видовой принадлежности по сохранившимся на своих местах остаткам полуободранных фотофоров — процедура, которая не терпит стандартного подхода. Определение поврежденных миктофид, на мой взгляд, имеет нечто общее с разгадыванием кроссворда, и поиск верного решения сам по себе очень увлекателен. Я люблю возиться с этими симпатичными рыбками, знаю многих из них «в лицо» и узнаю некоторых даже при полном отсутствии кожи, но тем не менее очень часто пасую перед непреодолимыми трудностями, доводя идентификацию только до родового уровня.
Светящиеся анчоусы очень широко распространены во всех широтах Мирового океана (их нет только в Центральном полярном бассейне), но достигают наибольшего видового разнообразия в тропических и субтропических водах. Все они принадлежат к числу океанических рыб, причем большинство видов постоянно обитают в толще воды и лишь немногие ведут бенто-пелагический образ жизни у материковых склонов и подводных поднятий. Среди пелагических миктофид резко преобладают среднеглубинные виды, которые держатся в светлое время суток на глубине порядка 300—1500 м. Для них очень характерны суточные вертикальные миграции с ночным подъемом на несколько сот метров в составе так называемых мигрирующих звукорассеивающих слоев (Звукорассеивающие слои - регистрируемые гидроакустическими приборами (эхолотами) слои, образованные скоплениями макропланктона,) (ЗРС) — комплекса макропланктонных и микронектонных животных, перемещающихся по вертикали в течение суток. В тропиках эти миграции, как правило, полностью укладываются в пределы основного термоклина (в высоких широтах верхняя граница сезонного слоя скачка также лимитирует выход миктофид к поверхности в летне-осеннее время). В то же время представители тепловодных родов Myctophum (кроме М. selenops), Symbolophorus, Gonichthys, Centrobranchus и Loweina (всего около 40 видов), относящихся к никтоэпипелагической группе, проникают ночью к самой поверхности воды. Батипелагические миктофиды (Taaningichthys, некоторые виды Lampanyctus и Lampadena) не меняют существенно горизонта своего обитания в течение суток.
Пелагические миктофиды — типичные макропланктон-ные рыбы, постоянно дрейфующие вместе с перемещающимися водами. Поэтому почти все видовые ареалы этих рыб находят объяснение в приуроченности их основ к тем или иным крупномасштабным круговоротам (из этого правила существуют, однако, отдельные не очень понятные исключения). Пассивным концентрированием особей определяются по-видимому, и скопления миктофид в некоторых районах схождения течений, например вдоль Южного полярного фронта.
Биология светящихся анчоусов изучена явно недостаточно. Известно, однако, что они почти безвыборочно питаются разнообразным мезопланктоном и принадлежат к числу консументов второго — третьего уровня. Продолжительность их жизни невелика: большинство тропических и мелкие умеренноводные виды живут всего 1—2 года, а более крупные умеренноводные —4—5 (по другим данным — 7—8) лет. В тропической зоне миктофиды размножаются круглогодично, а за ее пределами преимущественно в теплое время года, причем их личинки доминируют в эпипелагическом ихтиопланктоне всех районов открытого океана (это является косвенным доказательством их высокой численности). Некоторые тропические формы (в частности, никтоэпипелагические виды из родов Myctophum и Symbolophorus, а также Notolychnus valdiviae и др.) относятся к моноцикличным рыбам: они созревают уже в возрасте нескольких месяцев и после довольно продолжительного многопорционного нереста погибают примерно в годовалом возрасте. Индивидуальная плодовитость разных видов сильно варьирует, достигая нескольких тысяч икринок, и находится в прямой зависимости от размеров производителей.
Как правило, миктофиды держатся разреженными стайками, хотя в некоторых участках океана (например, в зоне антарктической конвергенции) они образуют более или менее значительные скопления. Такие скопления более характерны, однако, для отдельных участков над материковым склоном, и образуют их в основном специфические «присклоновые» виды. Для одного такого вида — Ceratoscopelus maderensis — у восточного побережья Северной Америки отмечены на глубине 600 м плотные линзовидные стаи диаметром до 100 м и толщиной до 10 м, в которых плотность рыб достигает 10—15 экз./м3 (в одной стае, следовательно, может насчитываться 300—400 тыс. особей средней длиной около 6 см). В целом светящиеся анчоусы, несомненно, являются наиболее многочисленной группой мезопелагических рыб, играющей огромную роль в пищевых цепях открытого океана.
К числу массовых рыб глубоководной пелагиали принадлежат также некоторые представители близкородственных семейств Photichthyidae (фотихтиевые) и Gonostomatidae (гоностомовые), которые, как и большинство других обитателей мезопелагиали, обладают хорошо развитыми органами свечения (рис. 18). Среди фотихтиевых особенно высокой численности достигают мезопелагичес-кие винцигуерии (род Vinciguerria), широко распространенные в тропических и субтропических водах. Впрочем, за исключением отдельных участков восточной части Тихого океана, где один из видов (V. lucetia) может доминировать в ихтиоцене, они нигде не конкурируют с миктофидами по биомассе. Пищу винцигуерий, максимальная длина которых не превышает 5,5 см, составляет мелкий планктон. Откорм происходит главным образом в верхних слоях океана (у V. nimbaria и V. lucetia иногда даже у поверхности), куда рыбы поднимаются ночью в составе ЗРС; горизонт их дневного обитания находится на глубине 300— 600 м. Продолжительность жизни не превышает 2 лет, причем начинающийся с наступлением половой зрелости примерно в годовалом возрасте порционный нерест продолжается до естественной гибели рыб.
Из гоностомовых рыб, в изучение которых большой вклад внесла В. А. Мухачева, необходимо в первую очередь отметить род циклотон (Cyclothone), который считается, если судить не по биомассе, а по числу особей, самым высокочисленным среди всех нынеживущих родов рыб. Сюда относятся 11 видов мелких рыбок (длина взрослых особей от 3 до 6—7 см, масса — от 60 до 600 мг - нижнее значение близко к минимальному для рыб), из из которых 2 вида мезопелагические, 6 — мезобатипелагические, 2 — батипелагические и 1 — батиабиссопелагический. Вертикальные миграции для циклотон не характерны. С увеличением глубины обитания окраска тела для видов меняется от прозрачной до чёрной, а размеры фотофоров уменьшаются и у самого глубинного вида отсутствуют. Светлые циклотоны, как правило, гораздо меньше тёмных по длине и биологически отличаются от них. У всех видов в 1,2-1,6 раза меньше самок и имеют сильно развитые обонятельные органы, служащие для отыскивания особей противоположного пола. Циклотоны питаются в основном мелкими планктонными ракообразными, причём батипелагические виды, живущие в бедных пищей водах, питаются от случая к случаю, даже не каждый день. Их рост и возраст почти не изучены(на отолитах C. acclinindens из северо-восточной Пацифики насчитывали не более 3 годовых колец). Плодовитость вартирует от 100 до 10 тыс икринок. Для двух батипелагических видов характерен протандрический гермафродитизм: все особи первоначально финкционируют как самцы, а затем большая их часть становится самками.

Рис. 18. Винцигерия (I), циклотона (3) и топорик аргиропелекус (3)
Такая же реверсия пола отмечена у тех видов близкородственного рода Gonostoma, которые обитают на большой глубине (G. bathyphilum, G. gracile). Гоностомы, как и циклотоны, принадлежат к числу планктофагов, но имеют более крупные размеры (от 7 до 27 см), и некоторые из них входят в состав мигрирующих ЗРС.
Очень своеобразным обликом отличается семейство рыб-топориков (Sternoptychidae) (в последние годы его изучали Р. Бэрд и О. Д. Бородулина), в котором, как и в двух рассматриваемых выше, объединяются как пелагические, так и бенто-пелагические виды (их длина варьирует от 4 до 10 см). Наиболее массовые мезопелагические роды (Argyropelecus и Sternoptyx) широко распространены в теплых водах. Топорики не совершают значительных суточных миграций: аргиропелекусы, характерную особенность которых составляют телескопические глаза, направленные вверх, держатся обычно в верхней мезопелагиали, на глубине 200—500 м, стерноптиксы — в нижней на глубине 500—1500 м. Возраст крупных особей A. pacificus длиной 6—7,5 см составляет 3—4 года. Пищей топориков служат в основном мелкие рачки, а сами они являются объектом питания многих хищников, включая тунцов, заходящих в пограничные с эпипе-лагиалью слои основного термоклина для откорма.
Еще одно семейство относительно некрупных (длина до 20 см) мирных рыб — батилаговые (Bathylagidae) представлено во всех зонах Мирового океана, но особенно характерно для умеренных и высоких широт. Достаточно сказать, например, что дальневосточная серебрянка (Leuroglossus schmidti) является самой массовой средне-глубинной рыбой Охотского моря. Батилагиды (в семействе, по С. Г. Кобылянскому, около 20 видов), подобно циклотонам, разделяются на две группы — «светлых» и «темных», преимущественно приуроченных в своем распространении соответственно к верхней и нижней мезопелагиали, причем некоторые светлые виды (тропический Bathylagoides nigrigenys) совершают вертикальные миграции в составе ЗРС. Батилаги питаются зоопланктоном, потребляя в основном не ракообразных, а животных с мягкими покровами — крылоногих моллюсков, щетинко-челюстных (хетогнат), аппендикулярий. По возрасту я росту данных очень мало. Известно, однако, что особи одного из темных северотихоокеанских видов при длине 11 см имеют 3, а при 15 см — 5 годовых колец на отолитах. Антарктический Bathylagus antarcticus нерестится 3—4 раза в году, выметывая до 5 тыс. икринок.
К семейству меламфаевых (Melamphaidae) относятся рыбы, населяющие толщу вод открытого океана и наиболее обычные на глубине 500—2000 м, т. е. в мезо- и батипелагиали. Они имеют однотонную темную окраску, большой рот с очень мелкими зубами и сильно развитую систему чувствительных к колебаниям воды сейсмосенсорных органов и слизевых каналов на голове. В состав семейства входит, по А. Эбелингу, около 35 видов, многие из которых имеют ограниченное распространение и приурочены к определенным водным массам. Интересно, что виды, населяющие наиболее кормные районы с высокой биологической продуктивностью, достигают больших размеров, чем те, ареалы которых приурочены к бедным пищевыми ресурсами водам. Так, в малокормных центральных круговоротах преобладают карликовые виды рода Melamphaes, длина которых во взрослом состоянии не превышает 2,5—3 см, тогда как размеры наиболее крупных представителей того же рода достигают 10—12 см. Пищу меламфаев составляют различные планктонные беспозвоночные, в основном мелкие ракообразные, а их самих поедают более крупные глубоководные хищники, в частности рыбы-удильщики. Несмотря на малые размеры тела, представители семейства имеют, по-видимому, довольно большую продолжительность жизни: поро-митры (род Poromitra), например, достигают 10-летнего возраста при максимальной длине 15 см. В отличие от многих других глубоководных рыб меламфаевые не совершают протяженных вертикальных миграций, хотя некоторые виды ночью несколько расширяют зону своего обитания.

Рис. 19. Глубоководные рыбы с телескопическими глазами: опистопрокт, или рыба-утюг (1), жемчужиноглазка (2), розаура (3), палочкохвоет (4)
Из других рыб, обитающих в глубоководной пелагиали, мелким планктоном питаются немногие и к тому же малочисленные преимущественно среднеглубинные виды, но некоторые из них несомненно заслуживают специального упоминания, хотя бы в связи со своим необычайным внешним обликом. Обладающий длинным, почти цилиндрическим в передней половине телом ксенофтальмихт (Xenophthalmichthys danae) имеет сильно увеличенные трубчатые глаза, направленные прямо вперед. Такие же телескопические глаза, но ориентированные вверх, развиты у видов семейства опистопроктовых (Opisthoproctidae), среди которых крайним своеобразием выделяются рыбы-утюги (два вида рода Opisthoproctus) (рис. 19). У этих небольших (до 10 см) рыбок брюшная поверхность тела образует совершенно плоскую «подошву», покрытую серебристыми чешуями и отражающую свет. У близкого рода Bathylychiiops с каждой стороны головы расположено по два глаза, из которых нормально развитый основной направлен косо вверх, а маленький дополнительный, находящийся под передней частью основного, — вперед и вниз. Этот дополнительный зрительный орган, развивающийся как вырост (дивертикул) стенки главного глазного яблока, имеет все структурные части глаза, включая хрусталик, и снабжен мускулатурой, позволяющей менять направление взгляда. Батилихнопс — уникальная четырехглазая рыба, которая пользуется преимуществами почти полного кругового обзора.
Хорошо развитые трубчатые глаза, смотрящие вперед, имеют и совсем не родственные всем этим рыбам гигантуровые (семейство Giganturidae с родами Gigantura и Rosaura), которые обитают на глубине 500—1500 м. Они обладают мощным! зубным вооружением и, несмотря на относительно мелкие размеры (до 17 см), питаются главным образом рыбами, заглатывая даже такую добычу, которая превышает их по величине (в желудке экземпляра Gigantura chuni длиной 8 см был однажды обнаружен хаулиод длиной 14 см). Личинки гигантуровых, подобно личинкам других глубоководных рыб, живут в эпипела-гиали. По внешнему виду они не имеют ничего общего с взрослыми особями и до последнего времени даже выделялись в особое семейство.
Еще один вид с телескопическими глазами — причудливый палочкохвост (Stylephorus chordatus) (длина до 28 см), живущий на глубине 300—800 м, имеет в нижней части хвостового плавника пару утолщенных и очень длинных лучей, образующих упругий прут, длина которого вдвое превышает длину тела; между этими лучами проходит канал боковой линии. Нормальное положение рыбы в толще воды — вертикальное, головой вверх (т. е. такое же, как у его родичей — других вогмеровидных рыб) — позволяет палочкохвосту пассивно дрейфовать, выжидая появления добычи — мелких копепод. Когда рачок оказывается поблизости, рыба резко выдвигает вперед свой трубчатый рот (ротовая полость при этом увеличивает объем почти в 40 раз) и втягивает в него жертву.
По-видимому, к числу мезопелагических рыб принадлежит и большеротая акула мегахазма (Megachasma pelagios) (рис. 20), известная пока лишь по двум экземплярам длиной 4,5 м. Первый из них был случайно пойман в 1976 г. у Гавайских островов — он запутался в плавучем якоре-парашюте научно-исследовательского судна на глубине 165 м, второй добыт сетями в 1984 г. в районе Калифорнии. В желудке этой акулы были найдены только эвфаузиевые раки тизаноподы, обитающие на глубине 150—1100 м. О глубоководном образе жизни этого вида свидетельствует также слабая мускулатура, мягкие покровы, малообызвествленные позвонки и другие особенности строения. Огромная пасть и хорошо развитые жаберные тычинки позволяют считать, что большеротая акула, подобно китовой и гигантской, является облигатным планктофагом-фильтратором, а серебристая выстилка нёба дала основание для предположения о том, что она может привлекать добычу, подсвечивая изнутри открытую пасть.
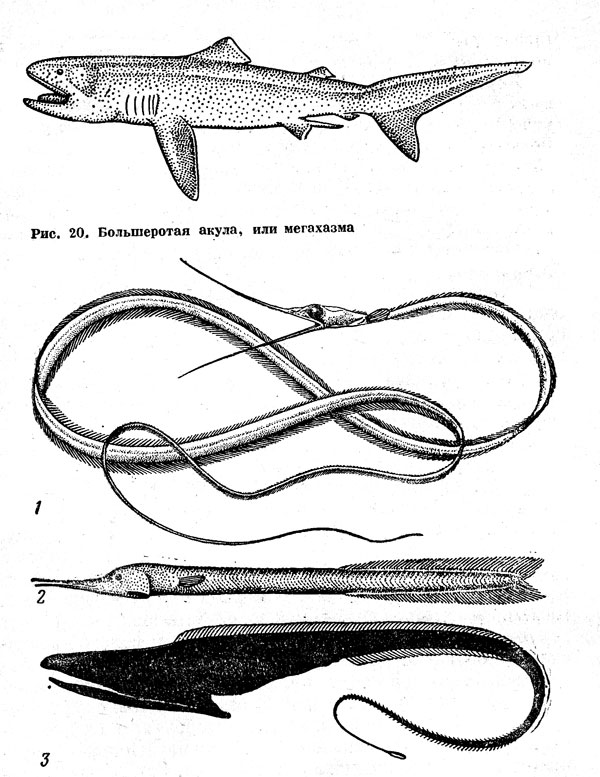
Рис. 20. Большеротая акула, или мегахазма. Рис. 21. Глубоководные пелагические угри: немихтис (1), циема (2), большерот, или пеликаний угорь (3)
Очень своеобразны и крайне редко встречающиеся мирапинновидные рыбы (подотряд Mirapinnoidei), но даже среди них особенно выделяется сенсационно знаменитая «волосатая», или «мохнатая», рыба Mirapinna esau, 30 лет назад часто упоминавшаяся в газетах разных стран. Этот вид до сих пор остается известным по единственному экземпляру длиной 4 см, пойманному у поверхности воды близ Азорских островов (по-видимому, случайный вынос с глубины). Тело рыбки, включая голову и плавники, действительно покрыто мягкими волосовидными выростами длиной до 1,5 мм (они несут железистые клетки, функция которых остается невыясненной). Представители близкого семейства Eutaeniophoridae, известные с глубины 300—1400 м, характеризуются наличием длинного лентообразного придатка на хвостовом плавнике.
Остальные мезо- и батипелагические рыбы питаются более крупными — макропланктонными организмами, в большинстве случаев не отдавая полного предпочтения каким-либо специфическим животным. Из этого правила есть, однако, несколько исключений, и все они связаны с выборочным потреблением ракообразных, главным образом эвфаузиид и креветок.
Среди таких стенофагов нужно отметить прежде всего нитехвостых угрей (Nemichthyidae), представленных как в верхней мезопелагиали (род Nemichthys), так и на больших глубинах — до 2000 м и глубже (род Avocettina) (рис. 21). Нитехвостые угри, достигающие 75—150 см в длину, характеризуются, как недавно выяснили И. Нильсен, Д. Смит и Э. С. Кармовская, очень интересными особенностями жизненного цикла. В период между ювенильным метаморфозом (эти рыбы, как и все угреобразные, имеют лептоцефальную личинку) и наступлением половой зрелости они обладают длинными тонкими челюстями, усаженными чрезвычайно мелкими частыми зубами, но созревшие самцы теряют и зубы, и челюсти, резко изменяя свой внешний облик (в связи с этим их даже выделяли в особые роды и виды). Есть предположения, что аналогичное превращение может происходить и с самками. Эти дегенеративные изменения, совпадающие с созреванием, и редкость поимок готовых к размножению особей позволяют думать, что нитехвостые угри гибнут сразу же после нереста. Продолжительность их жизни достигает 10 лет, а может быть, и большая. Из подводных аппаратов немихтиды наблюдались неподвижно зависшими в толще воды: по-видимому, таким способом они подкарауливают своих жертв — эвфаузиид и креветок сергистид, хватая их, когда те прикоснутся антеннами к щетке челюстных зубов и зацепятся за нее.
Для глубоководной пелагиали характерно еще несколько семейств угреобразных. Пилосошниковые угри (Serrivomeridae), длина которых достигает 75 см, довольно обычны в батипелагиали. Там же на глубине 1400—3000 м обитают и гораздо более редкие большероты, или пеликаньи угри (Eurypharynx pelecanoides), длиной до 1 м и мешкороты (9 видов рода Saccopharynx) длиной до 170 см, выделяющиеся непропорционально большой по сравнению с черепом пастью и сильно растяжимой глоткой (см. рис. 21). Еще глубже (до 5800 м) живут циемы (два вида родов Суета и Neocyema) — короткотелые плоские угри, экология которых почти не изучена.
Как ни удивительно, в составе мезопелагической фауны рыб находит себе законное место и обыкновенный европейский речной угорь, знакомый многим «сухопутным» рыболовам нашей страны, прежде всего, конечно, тем, кто ловит в реках Балтийского бассейна. Как-то и мне довелось присутствовать при поимке этой рыбы в одном из заливов Московского моря (оно же Иваньковское водохранилище). Местный парнишка с гордостью показал мне свою добычу: на дне лодки среди самых банальных окуньков я подлещиков извивался змеевидный полуметровый гость Подмосковья, явившийся сюда из весьма экзотических краев. Я сказал молодому рыбаку, что скользкая грязно-зеленая рыбинй принесла ему привет из далекого тропического океана, но не удивил его этим: будучи вполне современным человеком, он уже знал историю угря. Предложение моей сердобольной спутницы дать рыбе шанс вновь вернуться на свою родину - в теплое Саргассово море также было с ходу отвергнуто на основании полной нереальности такого возврата. Юный рационалист, бесспорно, был прав: вероятность такого события, действительно, бесконечно мала. Даже среди ученых есть скептики, которые сомневаются в возможности европейских угрей пересечь Атлантику. Но ведь угриное племя (а это как-никак особый вид!) все-таки существует, и, следовательно, кто-то все-таки возвращается на нерест, преодолевая все опасности на своем трудном пути и реализуя, казалось бы, совсем нереализуемые шансы...
Действительно, европейский угорь (Anguilla anguilla), как это было показано И. Шмидтом уже около 60 лет назад, размножается в глубинах центральной части Саргас-сова моря, воды которого отличаются наиболее высокой соленостью в пределах Северной Атлантики. Отсюда начинается дрейф икры, а затем личинок — лептоцефалов, пассивно переносимых на северо-восток поверхностными течениями. На это путешествие, которое заканчивается заходом прозрачных мальков (их называют стеклянными угрями) в европейские реки, уходит 2,5—3 года. В пресной воде самцы угря обычно проводят 6—12 лет, достигая длины 30—40 см, а самки —не менее 10—12 лет, вырастая за это время до 55—65 см. В океане взрослые угри мигрируют на запад, придерживаясь течений западного направления на глубине 1000—1200 м, достигают в конце концов места назначения и, отнерестившись там, погибают, дав начало новому поколению. Такая дальняя миграция требует, конечно, огромных энергетических затрат. На этом основании английский ихтиолог Д. Таккер предположил, что все взрослые особи, выросшие в реках Европы, гибнут в океане, не достигнув нерестилищ, а воспроизводство осуществляется за счет американской популяции (обычно она рассматривается в качестве особого вида A. rostrata). Различия между европейским и американским угрями этот автор относил на счет прямого влияния температуры в местах вымета икринок (в воды Европы попадают лептоцефалы, выклюнувшиеся в восточной части Саргассова моря) и отрицал, таким образом, само существование вида A. anguilla.
Перейдем теперь к хищным рыбам, пищу которых составляют объекты, сопоставимые с ними по размерам (не менее 15—20% длины хищника), — крупные ракообразные, головоногие моллюски, другие рыбы.(В ихтиологии под хищниками традиционно понимают рыб, питающихся другими рыбами, и противопоставляют их мирным рыбам, поедающим беспозвоночных животных и растения. В гидробиологии принято другое содержание термина: хищником считается любое животное, потребляющее в пищу животный корм ) В мезопелагиали наибольшего разнообразия среди них достигают представители «усатых стомиеобразных», или стомиевид-ных (сюда относятся 6 семейств), прежде всего мелано-стомиевые (Melanostomiidae), которых насчитывается, по современным представлениям, не менее 180 видов (рис, 22). Это — некрупные рыбы длиной от 10 до 40 см. Они имеют в той или иной степени удлиненное, лишенное чешуи тело интенсивного черного цвета, иногда отливающего металлическим блеском. У всех видов есть подбородочный усик, длина которого может у некоторых из них превышать длину тела. Система светящихся oрганов хорошо развита и включает обычно мелкие фотофоры, беспорядочно рассеянные по телу, крупные туловищные фотофоры, расположенные в продольных рядах вдоль нижнего профиля, большие заглазничные фотофоры, а также светящиеся органы на усике и пятна люминесцентной ткани на голове п туловище. Аквариальные наблюдения над эхиостомой (Echiostoma barbatum) показали, что свет, испускаемый заглазничным органом, может меняться от ярко-розового до зеленовато-белого. Цвет крупных фотофоров у особей этого вида варьирует от красноватого до темно-алого, мелких фотофоров — от розового до пурпурного, а светящаяся ткань на теле имеет зеленоватый цвет. Исключительного разнообразия (как по цвету, так и по числу, форме п расположению) достигают светящиеся железы на подбородочном усике, строение которого у многих видов рода Eustomias (всего их более 100) служит, как установлено Р. Гиббсом, единственным строгим признаком для их идентификации. Ме-ланостомиевые довольно обычны в тропических и субтропических водах, многие живут в умеренных широтах. В дневное время они концентрируются на глубине 500— 1500 м, а ночью поднимаются к верхней границе основного термоклина (молодые экземпляры Photonectes albipennis даже к поверхности воды). Питаются преимущественно рыбами (светящимися анчоусами, циклотонами), но некоторые виды (например, северотихоокеанская Tactostoma macropus) отдают предпочтение крупным ракообразным. Предельный возраст — 8 лет, а возможно, и больше.
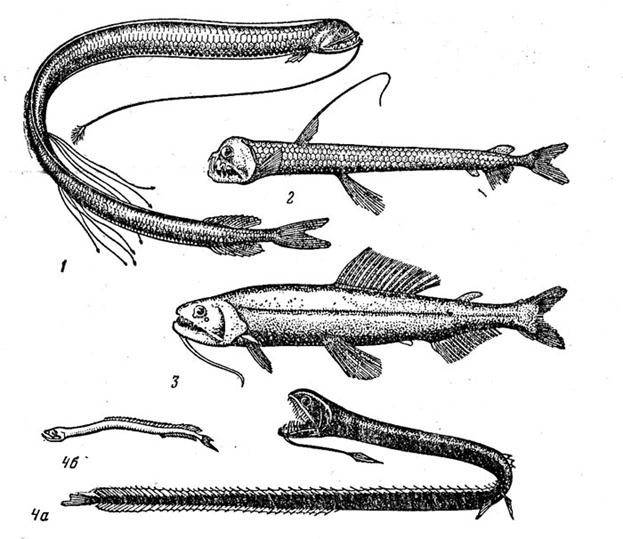
Рис. 22. Стомиеобразные рыбы: стомия (1), хаулпод (8), астронестес (3), идиакант — самка (4а) а самец (4Б)
В течение нескольких лет - это было в середине 70-х годов — меланостомиевые рыбы (на профессиональном жаргоне их называют «черненькими») находились в самом центре моих научных интересов. Представлялось поразительным, что в этой исключительно многообразной группе виды многих родов (например, эустомиасы и меланостомиасы) различаются между собой практически только по строению чрезвычайно сложно устроенного подбородочного усика, и хотелось разобраться в изменчивости этого аппарата. Надо сказать, что изучение этих усиков, несущих миниатюрные светящиеся луковицы и разного рода придатки,— исключительно тонкое дело, и мне очень повезло в том отношении, что я смог кооперироваться в этой работе с художницей нашей лаборатории Глафирой Похильской. Ведь в данном случае никакие словесные описания не могут заменить в высшей степени информативного научного рисунка.
Обработка собранной в Тихом и Индийском океанах коллекций дала возможность описать несколько новых форм и высказать предположения об эволюционных тенденциях в пределах группы. Наш материал оказался, однако, не очень представительным, и это исследование не получило дальнейшего развития. Более обширные коллекции эустомиасов были изучены американскими коллегами, которые пришли к заключению об исключительно высокой видоспецифичности строения усика и, основываясь главным образом на этой особенности, чуть ли не удвоили число видов в роде. Их результаты значительно продвинули вперед изучение этих своеобразных рыб, однако я думаю, что внутривидовая изменчивость усиков в ряде случаев осталась все-таки недооцененной, и именно этим объясняются странные узколокальные ареалы некоторых видов в Атлантическом океане. В общем, до полной ясности в познании видового состава «черненьких» еще далеко.
Близкие к мелоностомиевым семейства астронестовых (Astronesthidae) с 5 родами (35 видов) и монотипические хаулиодовые (Chauliodontidae, 8 видов), стомиевые (Stomiatidae, 14 видов), идиакантовые (Idiacanthidae, 3 вида) также имеют приспособления к питанию относительно крупной добычей — очень большой рот, острые клыковидные зубы, откидывающуюся вверх при глотании голову, огромный желудок (см. рис. 22). Все они используют при охоте сходную тактику — выжидание и приманивание жертв, — и все способны заглатывать добычу, лишь в 2—3 раза уступающую им по размерам. Пассивный образ жизни дает полное основание относить стомиевидных рыб к макропланктонной группировке.
Пожалуй, наиболее соответствует представлениям о страшных глубоководных хищниках — недаром они так часто изображаются в популярных книгах — хаулиоды с их гигантскими кривыми клыками, которые не умещаются в пасти и устрашающе торчат наружу даже при закрытом рте (эти страшилища, правда, невелики по размеру: самый крупный вид Chauliodus sloani не превышает 30 см в длину). Подбородочный усик у всех хаулиодов очень маленький, но первый луч спинного плавника вытянут в длинную нить со светящимся кончиком, которая, как показывают подводные наблюдения, свободно свисает перед ртом и служит для привлечения доверчивых жертв.
Идиаканты представляют интерес прежде всего с точки зрения их биологии. Самки, имеющие длинное угревидное тело, достигают длины 40 см и по образу жизни не отличаются от других стомиевидных рыб. Этого никак нельзя сказать о самцах — они гораздо мельче (длина всего лишь 6—7 см) и отличаются от самок отсутствием зубов, усика, брюшных плавников и, главное, полной дегенерацией пищеварительного тракта. По окончании личиночной стадии (личинки идиакантов, кстати сказать, очень своеобразны — их глаза сидят на концах длиннейших, тонких, как нитка, стебельков, что облегчает парение в воде) самцы не питаются и вряд ли живут дольше нескольких месяцев. В то же время самка северотихоокеанского вида Idiacanthus antrostomus длиной 32,5 см имела 6 лет от роду. Плодовитость крупной самки тропического вида I. fasciola определена в 14 тыс. икринок.
Еще одна весьма примечательная группа макропланктонных рыб — церациевидные или глубоководные удильщики (подотряд Ceratioidei), очень большой вклад в изучение которых внесен Э. Бертельсоном и Т. Питчем, весьма характерна для бати- и абиссопелагиали. Сюда относятся 11 семейств и около 120 видов, довольно разнообразных по размерам и внешнему облику (рис. 23), но обладающих одной общей особенностью — видоизмененным в «удилище» (иллиций) первым лучом спинного плавника. На конце иллиция находится «приманка» (эска), которая у разных видов сильно различается по строению, величине и форме. Все удильщики имеют довольно короткое тело (исключение составляет семейство Gigantactinidae) с непропорционально большой головой, огромный, как правило, зубастый рот и сильно растяжимый желудок. Окраска однотонная черная, реже темно-коричневая. Органов свечения на туловище и голове нет, и у большинства видов единственной светоизлучающей структурой служит железа, входящая в состав эски. Исключение составляют только линофрины (род Linophryne), у которых в дополнение к удочке имеются подбородочные усики со светящимися придатками. В заполненном слизью просвете эскальной трубчатой железы живут светящиеся симбиотические бактерии, которых до сих пор не удалось размножить в культуре и изучить детально. Эскальная железа и ее придатки (иногда сложно устроенные) изолированы от наружной среды отражающим и пигментным слоями, прободаемыми «окнами» и «щелями», которые обеспечивают видоспецифичность иллюминации эски. Регуляция интенсивности свечения обеспечивается только изменением интенсивности кровоснабжения, т. е. скорости подачи в железу кислорода, необходимого для жизнедеятельности бактерий. Функциональная роль эски как приманки, привлекающей кормовые объекты удильщиков, совершенно очевидна, однако серьезные межвидовые различия в строении органа заставляют предполагать и его коммуникационное значение.