На примере аборигенов Австралии и Океании 7 страница

Рис. 11. Номей: малек (рыба-пастушок) под сифонофорой физалией (1) и взрослый экземпляр(2).
Особенно большую известность благодаря облигатном комменсализму молоди с плавающими на поверхности сифонофорами физалиями, яд которых опасен даже для человека, получил номей (Nomeus gronovii) (рис. 11), распространенный в теплых водах всех океанов. Пестрые мальки этого вида, имеющие яркие темно-синие пятна на серебристом теле, как правило, не встречаются вдали от сифонофор и получили в связи с этой особенностью поведения название «рыбы-пастушки». Пища физалий состоит из молоди рыб, которую они убивают стрекательными клетками, расположенными на ловчих шупальцах. Но маленькие номей относительно иммунны к токсину физалий, хотя и стараются при этом избегать непосредственного контакта с их щупальцами (известны, впрочем, и отдельные случаи поедания рыбок их опасными «хозяевами»). Несмотря на то что под сифонофорами отмечались лишь мальки и неполовозрелые особи длиной от 1 до 15 см, долгое время считалось, что сожительство номея и физалий имеет постоянный в течение всей жизни характер. Недавно было показано, однако, что взрослые рыбы этого вида живут самостоятельной жизнью, о чем свидетельствуют поимки большим пелагическим тралом экземпляров длиной до 35 см, резко отличающихся от более мелких однотонно-бурой окраской и совсем другими пропорциями тела.
Кубоглавы (род Cubiceps с 10 видами), обладающие очень длинными грудными плавниками, также обычны во всех районах, тропической и субтропической зон. Распространенный в Северо-Восточной Атлантике обыкновенный кубоглав (С. gracilis) в раннем возрасте держится у поверхности и часто ловится под медузами. Половое созревание наступает при достижении длины 20 см, но и после этого рост рыбы продолжается и максимальная длина достигает 1 м. Такие крупные особи живут в подповерхностных слоях. Другие виды кубоглавов значительно мельче: многие из них, например С. pauciradiatus, не превышают 20—25 см.
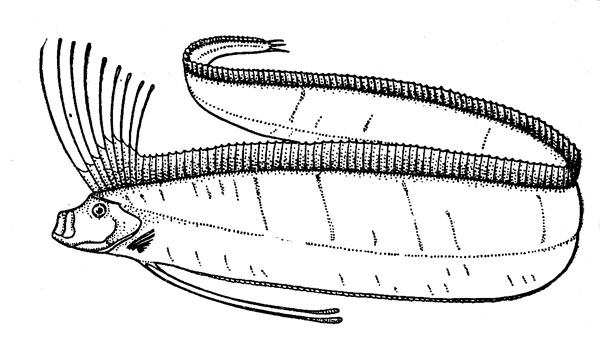
Рис. 12. Сельдяной король.
Некоторые представители рассматриваемого подотряда, например центролоф (Centrolophus niger), живущий в умеренно теплых водах Северной Атлантики и южного полушария, по мере роста существенно меняют глубину обитания. Мальки этого вида держатся у поверхности и, подобно другим строматеевидным, сопровождают крупных медуз. Они быстро растут (в аквариуме рыбка длиной 2 см всего за пять месяцев увеличила размер в 8,5 раз). Взрослые центролофы — однотонные черно-коричневые рыбы, длина которых достигает 1,2 м, — держатся только в нижних горизонтах эпипелагиали.
Для нижней эпипелагиали характерны и другие «полуглубоководные» рыбы, обитающие также в верхней мезопелагиали. В их числе следует упомянуть прежде всего представителей подотряда вогмеровидных (Trachipteroidei), включающего 9 родов (около 15 видов), в том числе сельдяных королей (Regalecus glesne) (рис. 12) и вогмеров (род Trachipterus). Для этих рыб характерен аллометрический рост, т. е. большая онтогенетическая изменчивость формы тела и относительных размеров его частей, и пот этому мальки, молодь и взрослые особи некоторых видов долго носили разные научные названия. Вогмеровидныв рыбы имеют удлиненное и плоское — ремнеобразное — тело, сужающееся к хвосту, и отличаются большой длиной (не менее 1м). Сельдяной король достигает в Северной Атлантике даже длины 5,5 м при массе порядка 250 кг (известны и случаи поимки рыб длиной 7—8 м), а лососевый король (Trachipterus altivelis) из северо-восточной части Тихого океана в возрасте 7—8 лет имеет длину 1,6 м и весит 4 кг. (При сходной в общих чертах форме тела соотношения этих цифр кажутся довольно странными, но они вполне соответствуют представлениям о пропорциональности массы рыб кубу их длины.)
Вогмеровидные — хищники, питающиеся разнообразными рыбами и беспозвоночными. Иногда они встречаются в косяках массовых промысловых видов, в частности сельди и дальневосточных лососей, которых, как считали в давние времена, ведут за собой (отсюда и название «короли», но на деле их связи с «подданными» имеют чисто гастрономический характер). Эти рыбы плавают обычно головой кверху, в положении, близком к вертикальному. При этом они поддерживают от опускания собственное тело, плотность которого больше, чем плотность воды, и поступательно перемещаются с небольшой скоростью за счет одних только ундулирующих движений длинного спинного плавника. Если нужно плыть быстрее, короли передвигаются, волнообразно изгибая свое длинное тело. У всех представителей подотряда тело окрашено в серебристо-белый цвет, на туловище, как правило, разбросаны короткие темные полосы и пятна, плавники у многих ярко-красные. Виды семейства лофотовых (Lophotidae) имеют своеобразный чернильный мешок, открывающийся в клоаку, и способны создавать маскирующую «дымовую завесу» подобно кальмарам и каракатицам.
Вероятно, встречи моряков с гигантскими сельдяными королями, плавающими у поверхности, были причиной многих историй о «морском змее», который в некоторых рассказах описывается как чудовище, имеющее лошадиную голову с развевающейся огненно-рыжей гривой. За такую гриву, видимо, принимали длинные лучи спинного плавника, образующие головной султан этой рыбы.
Еще одно нижнеэпипелагическое (или эпимезопелагическое) семейство — морские лещи (Bramidae) — также приурочено преимущественно к субтропическим и тропическим водам, хотя отдельные виды обитают и в сравнительно холодной зоне. Атлантический лещ (Вгата Ьгата), например, населяет Средиземное море и северную Атлантику и встречается при температуре 10—15°. В южной части области распространения он живет на глубине, а у его северных границ обычен и в поверхностном слое. Эта хищная рыба достигает длины 65—70 см и массы 6 кг и имеет, как и близкородственный северотихоокеанский лещ; (В. japonicus), некоторое промысловое значение. Среди более редких видов можно отметить серебряного морского леща (Pterycombus brama), рыбу с блестящим серебристым телом, желтыми грудными и иссиня-черными спинными а анальными плавниками; при этом оба вертикальных плавника сильно увеличены и имеют парусовидпую форму.
Рассматривая нижнеэпипелагических рыб, нельзя ни упомянуть еще один интересный вид — луваря (Luvarua imperialis), которого выделяют в особое семейство Luvaridae. Это семейство длительное время относили к скумбрие видным рыбам, но недавно было показано, что оно наиболее родственно, как это ни удивительно, прибрежным рыбам-хирургам (семейство Acanthuridae), обитающим главным образом на коралловых рифах. Луварь имеет удлиненное плоское тело и лобастую голову с крохотным ротиком. Он достигает 2 м в длину и весит до 150 кг, очень плодовит (47 млн ооцитов), его личиночные и мальковые стадии очень мало похожи на взрослых луварей. Основной пищей являются медузы и другие студенистые формы планктона, т. е. низкокалорийные объекты, и в связи с этим, как считают Д. Фитч и Р. Лавенберг, он обладает очень длинным кишечником (у метровой рыбы он составлял 92% длины тела). Луварь распространен во всех океанах, но всюду редок, и большинство поимок приходится на субтропическую зону.
Совершенно особую и очень своеобразную группировку в составе эпипелагического ихтиоцена представляют рыбы, ассоциированные с плавучими водорослями саргассами, которые обычно выносятся в океан с прибрежных мелководий, но кое-где (например, в Саргассовом море) существуют в замкнутых круговоротах без всякой связи с берегами. Помимо молоди некоторых океанических рыб (летучки, корифены) и рыб прибрежного происхождения, среди саргассов постоянно обитают два высокоспециализированных вида — морской клоун Histrio histrio (длина до 18 см) и пелагическая рыба-игла Syngnathus pelagicus (длина до 10 см). Внешний вид и окраска этих рыб, никогда не выходящих за пределы водорослевых кустов и скоплений (рис. 13), отлично маскируют их в саргассах, и их нелегко, обнаружить в этом укрытии, даже точно зная, что они скрываются там. Рыба-клоун привлекает добычу специальной приманкой своей «удочки» (первого луча спинного плавника) и заглатывает ее, быстро увеличивая объем ротовой полости при закрытом рте и открывая пасть, когда жертва оказывается прямо перед ней, — весь этот процесс укладывается в тысячные доли секунды.

Рис. 13. Рыбы, ассоциированные с плавучими водорослями.
Почти все рыбы, перечисленные выше, живут в эпипелагиали постоянно — со времени появления на свет до конца своего существования. Таких рыб предложено называть голоэпипелагическими и противопоставлять их видам меропипелагическим, в обязательном порядке проводящим в верхних горизонтах открытого океана лишь определенную часть своего жизненного цикла, и ксеноэпипелагическим, представляющим собой более или менее случайный элемент в поверхностном слое (см. рис. 14) ( «Голос» по-гречески - весь, целый, «мерос» - часть, доля, «ксенос» - чуждый.).

В 1964 г. директор Института океанологии В. Г. Корт поставил перед сотрудниками задачу обобщить в коллективной монографии все сведения о природе Тихого океана, накопленные к тому времени. Мне было поручено свести воедино то, что было известно об эпипелагических рыбах, которые тогда изучались советскими экспедициями (напомню, что в дальнем океане в те времена фактически работал один «Витязь») в очень узких рамках — в открытых водах проводились только световые станции и сборы ихтиопланктона. В отношении наиболее массовых и соответственно наиболее интересных для промысла видов - тунцов, марлинов и других - пришлось использовать только литературные данные. Эта компилятивная работа приносила мне мало удовлетворения (особенно на ее начальном этапе), но именно она заложила основы моих представлений о сложной структуре «композитного» океанического ихтиоцена. После выхода в свет многотомника «Тихий океан», который получил высокую оценку (группе его авторов была присуждена Государственная премия СССР), я значительно расширил свои главы, дополнил их сведениями по другим океанам и. подготовил монографию «Ихтиофауна океанской эпипелагиали», которая стала моей докторской диссертацией. Ее основные положения вошли в эту книгу но, конечно, в значительно видоизмененном изложении: за 20 лет, прошедших со времени ее опубликования, изучение рыб открытого океана очень сильно продвинулось вперед, да и сам я стал по-иному смотреть на многие вещи. Сейчас, например, я никак не дал бы той книжке ее старого названия (ведь в ней речь идет не столько о «фауне», сколько о «таксоцене»), отказался бы от ненужного, как вижу теперь, придумывания новых, порой труднопроизносимых терминов для обозначения отдельных экологических группировок рыб, внес бы многочисленные коррективы в биологические характеристики отдельных видов. Впрочем, «что написано пером, того не вырубишь топором»...
Итак, по числу видов временные обитатели эпипелагиали не уступают голоэпипелагической группе. Некоторые из них достигают в отдельных районах океана очень высокой численности и биомассы.
Все мероэпипелагические рыбы используют эпипелагиаль посредством замкнутых миграционных циклов. Именно этой особенностью они достаточно четко отличаются от ксеноэпипелагических рыб, не совершающих регулярных и целенаправленных перемещений между эпипелагиалыо ц другими биотопами. Большинство ксеноэпипелагических видов нормально обитают в неритической зоне, но, попав за ее пределы, они способны долго существовать (по-видимому, не размножаясь) и в практически неограниченном удалении от берегов. В качестве примера можно упомянуть хотя бы представителей трех семейств отряда сарганообразных — парусноперую летучую рыбу Parexocoetus brachypterus (семейство Exocoetidae), длинноносого полурыла Euleptoramphus viridis (Hemiramphidae), лентовидного саргана Ablennes hians (Belonidae), которые, являясь по своему существу неритическими, могут быть встречены в любой точке тропического океана. В северной части Тихого океана, в Баренцевом и Черном морях на большом расстоянии от берега можно наблюдать трехиглую колюшку, или колючку (Gasterosteus aculeatus),— обычного обитателя наших рек, озер и прибрежных морских вод. К этой же группе следует отнести и тех прибрежных альгофильных рыб, которые встречаются в тропической эпипелагиали вместе с плавающими водорослями или другими дрейфующими предметами берегового происхождения, например крокодилового саргана (Tylosurus crocodilus), пятнистого спинорога (Canthidermis maculatus), абудефдуфа (Abudef-duf saxatilis), терапона (Therapon theraps), чаба (Kypho-sus cinerascens). Как показал Б. И. Федоряко, эти и некоторые другие рыбы, будучи вынесенными в открытые воды еще на мальковой стадии, прекрасно чувствуют себя и вдали от берегов, там, где преобладающее большинство прибрежных рыб гибнет очень быстро. Способность к долгому выживанию в эпипелагиали, несомненно, содействует широкому расселению ксеноэпипелагических видов, для которых не существует барьеров в виде открытоводных пространств любого протяжения. Многие из них имеют поэтому циркумглобальные ареалы.
Временные обитатели эпипелагиали проникают в нее либо снизу — из глубинных вод, либо «сбоку» — из неритической зоны и с прибрежных мелководий. При этом мероэпипелагические глубоководные рыбы появляются в эпипелагиали лишь ночью (здесь речь идет только о взрослых особях), когда опи поднимаются вверх в ходе регулярных суточных миграций, а перед рассветом вновь уходят на глубины. К этой никтоэпипелагической группе принадлежат уже упоминавшиеся мельком приповерхностные светящиеся анчоусы и змеиные макрели, а также некоторые карликовые акулы и отдельные представители типичных мезопелагических семейств, о которых пойдет речь в следующей главе. Следует отметить, однако, что в отсутствие резкого температурного градиента между приповерхностными и глубинными водами (это характерно для внетропической зоны) обособленность эпи- и мезопелагической группировок рыб выражена значительно слабее, чем в районах с хорошо развитым термоклином, представляющим собой препятствие для вертикальных мигрантов.
Мероэпипелагические рыбы прибрежного происхождения, которые встречаются в открытых водах только в период нагула, по размножаются у берегов или даже в пресных водах, составляют очень характерный элемент населения эпипелагиали. Они нередко образуют значительную биомассу, в некоторых районах соизмеримую с биомассой постоянных обитателей эпипелагиали или даже превышающую ее. Типичными представителями этой группировки являются, в частности, настоящие (род Salmo) и тихоокеанские (Oncorhinchus) лососи. Представители последнего рода — кета, горбуша, нерка, кижуч, чавыча, сима — эндемичны для бассейна северной части Тихого океана и в период нагула являются наиболее массовыми рыбами поверхностного слоя на огромной акватории — от Берингова моря до 40° с. ш. Тихоокеанские лососи образуют многочисленные репродуктивные стада, нерестящиеся в реках дальневосточного побережья Азии (на юг до п-ова Корея и Японии; жилая форма нерки есть даже на о-ве Тайвань) и Северной Америки (до Калифорнии). Ареалы таких стад в открытом океане в значительной степени перекрываются, и между 165° в. д. и 155° з. д. даже азиатские и американские лососи встречаются совместно. Длительность морского периода жизни у разных видов неодинакова — от 1,5 лет у горбуши до 4—7 лет у чавычи, наиболее крупного вида, иногда достигающего 50 кг. Они питаются среднеразмерными и крупными планктонными животными — ракообразными, крылоногими моллюсками, молодью кальмаров и рыб. Границы морских ареалов тихоокеанских лососей претерпевают значительные сезонные изменения, однако даже в зимний период область их распространения не простирается за пределы субарктических вод.
Атлантическо-скандинавская (так называемая океаническая) сельдь (Clupea harengus) размножается в прибрежных водах Норвегии и Исландии, откладывая клейкую икру на водоросли, а в период нагула ведет пелагический образ жизни вдали от берега. В годы хорошего состояния запасов она совершает довольно протяженные миграции в открытую часть Северной Атлантики и достигает Баренцева моря, Шпицбергена, Гренландии (сейчас численность этого вида находится в состоянии депрессии). Примерно в этом же районе в современный период происходит нагул северной путассу (Micromesistius poutassou) — стайной рыбы семейства тресковых (Gadidae), размножение которой также ограничено склонами мелководий в неритической зоне.
Самая большая из хищных рыб Южного океана — антарктический клыкач (Dissostichus mawsoni) размножается у берегов Антарктиды, где держится и подрастающая молодь длиной до 1 м, тогда как более крупные (до 1,8 м) половозрелые рыбы, как было показано В. Л. Юховым, совершают долговременные миграции в пелагиаль, удаляясь па расстояние до 900 миль от материка. Такое распространение клыкачей было установлено по их нахождениям в желудках кашалотов, питающихся преимущественно на глубине 100—1000 м, что дало основание относить эту рыбу к временным обитателям мезопелагиали. Может быть, это действительно так, но, учитывая недостаточность имеющихся данных, нет оснований признавать строго обоснованным тезис об отсутствии клыкача в поверхностных горизонтах.
Другая группа мероэпипелагических видов представлена в эпипелагиали только ранними стадиями развития (вплоть до мальков). Для всех этих видов, живущих во взрослом состоянии в неритической пелагиали или у дна, прохождение эпипелагической стадии в онтогенезе является необходимой и неизбежной фазой жизненного цикла. Будучи хорошо адаптированными к весьма длительному пребыванию в толще воды, такие мальки нередко встречаются в огромном удалении от берегов и сохраняют шансы на благополучное возвращение в прибрежную зону, хотя значительная их часть, конечно, обречена на неизбежную гибель в открытых водах.
Разделение эпипелагических рыб открытого океана между голо-, меро- и ксеноэпипеелагической группировками, как правило, достаточно очевидно, хотя не абсолютно — как обычно, желание разложить природное разнообразие по полочкам вступает в противоречие с практикой жизни. Так, многие виды крупных акул (в частности, кархародон и тигровая), большая барракуда (Sphyrae-na barracuda), ауксиды, полосатый и желтоперый тунцы, ваху, большая корифена, по-видимому, в той или иной степени обычны и в неритической зоне, и в открытых водах (часто их называют нерито-океаническими). Не во всех случаях при современной степени экологической изученности можно четко разграничить мероэпипелагических и ксеноэпипелагических рыб.
Полный перечень отрядов, семейств и родов, представленных в эпипелагиали открытого океана, приводится ниже. Для каждого из родов указаны в скобках число эпипелагических видов и их принадлежность к числу постоянных (Г), временных (М) и случайных (К) обитателей биотопа (виды, представленные в эпипелагиали только молодью, не учтены).
Систематический состав эпипелагического ихтиоцена
Отряд миногообразные (Petromyzontiformes). Сем. миноговые (Petromyzontidae): Geotria (1 К).
Отряд катранообразные (Squaliformes). Сем. катрановые (Squalidae): Euprotomicrus (1 М); к числу мигрирующих ночью из мезопелагиали или живущих на стыке мезо- и эпипелагиали относятся еще 9 видов из 6 родов — Isistius, Squaliolus, Euprotomicroides и др. Отряд воббегонгообразные (Orectolobiformes). Сем. китовые акулы (Rhiniodontidae): Rhmiodon (1 Г).
Отряд ламнообразные (Lamniformes). Сем. ложнопесчаные акулы (Pseudocarchariidae): Pseudocarcharias (1 Г); лисьи акулы (Alopiidae): Alopias (1 Г и 2 К); большеротые акулы (Megachasmidae): Megachasma (1 К); гигантские акулы (Cetorhinidae): Cetorhinus (1 К); сельдевые акулы (Lamnidae): Lamna (2 Г); Isurus (2 Г), Carcharodon (1 К). Отряд кархаринообразные (Carcharhiniformes). Сем. серые акулы (Carcharhinidae): Carcharhinus (2 Г и 5-6 К), Prionace (1 Г), Galoocerdo (IK); молотоголовые акулы (Sphyrnidae): Sphyrna (3 К). Отряд хвостоколообразные (Myliobatiformes). Сем. скаты-хвостоколы (D аsyatidae): Dasyatis (1 Г); мантовые скаты (Моbulidaa): Mania (1 К).
Отряд гнюсообразиые (Torpediniformes). Сем. гнюсовые (Torpedinidae): Torpedo (2-3 К).
Отряд угреобразные (Anguilliformes): Сем. змеехвостые угри (Ophichthyidae): Benthenchelys (1 Г). Отряд сельдеобразные (Clupeiformes). Сем. сельдевые (Clupeidae): Clupea (1 M), Sardinops (1 К); анчоусовые (Еngraulidae); Stolephorus (1 Г), Engraulis (1 К). Отряд лососеобразиые (Salmoniformes). Сем. лососевые (Salmonidae): Oncorhynchus (5 M), Salmo (3 M). Отряд стомиеобразные (Stomiiformes). Сем. фотихтовые (Photichthyidae): Vinciguerria (2 M); астронестовые (Astronesthidae): Astronesthes (1-2 M). Отряд аулопообрэзиые (Aulopiformes). Сем. алепиэавровые (Alepisauridae): Alepisaurus (2 К); кинжалозубовые (Anotopteridae): Anotopterus (1 К), веретенниковые (Paralepididae): Paralepis и др. (3—4 К). Отряд миктофообразные (Myctophi formes). Сем. миктофовые (Myctophidae): Electrona (2 M), Benthosema (1 Г), Hygophum (2 M), Myctophum (13 M), Symbolophorus (около 10 M), Loweina (4 M), Tarletonbeania (1 M), Gonichthys (4 M), Centrobranchus (4 M). Отряд трескообразиые (Gadiformes). Сем. тресковые (Gadidae): Micromesistius (2 МЭн).
Отряд удильщикообразные (Lophiiformes), Сем. рыбы-клоуны (Antennariidae): Histrio (1 Г). Отряд capганообразные (Beloniforraes). Сем. летучие рыбы (Ехосоеtidae): Parexocoetus (1 К), Exocoetus (3 Г), Prognichthys (4 Г и 1 К), Cypselurus (5-6 К), Cheilopogon (5 Г и 10-12 К), Hirundichthys (3 Г и 3 КЭи);полурыловые (Hemiramphidae): Oxyporhamphus (2 Г), Euleptorhamphus (2 К); сарга новые (Belonidae): Platybelone (1 К), Ablennes (1 К), Tylosurus (2 К); макрелещуковые (Scomberesocidae): Scomberesox (1 Г), Cololabis (1 Г), Nanichthys (1 Г), Elassichthys (1 Г). Отряд опахообразные (Lampridiformes). Сем. опаховые (Lampridae): Lampris (2 Г); лофотовые (Lophotidae): Lophotus (1 Г), Eumecichthys (1 Г); радицефаловые (Radiicephalidae): Radiicephalus (1 Г); вогмеровые (Тrасhiptоridae): Trachipterus (5Г), Desmodema (2 Г), Zu (2Г); ремень-рыбы (Regalecidae): Regalecus (1 Г), Agrostichthys (1 Г).
Отряд колюшкообразные (Gasterosteiformes). Сем. колюшковые (Gasterosteidae): Gasterosteus (1 К); рыбы-иглы (Syngnathidae): Syngnathus (1 Г). Отряд екорпенообразмме (Scorpaeniformes). Сем. пипагоровые (Сус1орtегidае): Pelagocyclus (1 Г), Cyclopterus (1 К). Отряд окунеобразные (Pcrciformes). Сем. ставридовые (Carangidae): Elagatis (1 Г), Naucratus (1 Г), Decapterus (2-3 К), Trachurus (2-3 К); рыбы-прилипалы (Есhеnеidае): Phtheiricbthys (1 Г), Remoга (5 Г), Remorina (1 Г); корифеновые (Соrурhaenidae): Coryphaena (2 Г); морские лещи (В га mid а с): Таrасtes (2 Г), Brama (6 Г), Taractichthys (2 Г), Pterycombus (2 Г), Ptoraclis (3 Г); нототениевые (Nototheniidae): Dissostichus (2 M), Pleuragrainma (1 К), Paranotothenia (1 К); и костеевые (Icosteidae): Icosteus (1 Г); лунарепые (Luvari- Jae): Luvarus (1 Г); барракудовые (Sрhуrnidae): Sphyrna (1 К); темнило вые (Gеmруlidae): Lepidocybium (1 К). Ruvettus (1 К), Gempylus (I M); скумбриевые (Sсоmbr idae): Gasterochisma (1 Г), Scomber (2 К), Acanthocybium (1 ГЭв), Allolhunnus (1 Г), Auxis (2 Г), Euthynnus (2 КЭп), Katsuwonus (1 Г), Thunnus (7 Г); меч-рыбы (Xiphiidae): Xiphias (1 Г); парусников ые (Istiophoridae): Istiophorus (1 К), Makaira (3 Г), Tetrapterus (5 Г); центролофовые (Сentrolophidae): Centrolophus (1 Г), Icichthys (1 Г), Pseudoicichthys (1 Г), Tubbia (1 Г), Schedophilus (5 Г); амарсиповыэ (Amar sipida e): Amarsipus (1 Г); номе ев ые (Nоmоidae): Cubiceps (9 Г), Nomeus (1 Г), Psenes (6 Г); алетовые (Tetragonuridae): Tetragonurus (3 Г). Отряд скалозубообразныс (Tetraodontif ormes). Сем. спи пороговые (Balistidae): Canthidermis (1 Г), Balistes (1 К), Xanthichthys (2 К); ска л о зубовые (Tetraodontidae): Lagocephalus (1 Г); ежи-рыбы (Diodontidae): Diodon (1 Г + 2 К); рыбы-луны (Мо1idae): Ranzania (1 Г), Masturus (1 Г), Mola (2 Г).
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пе-лагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спиноорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus (рис. 15). Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Fix выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
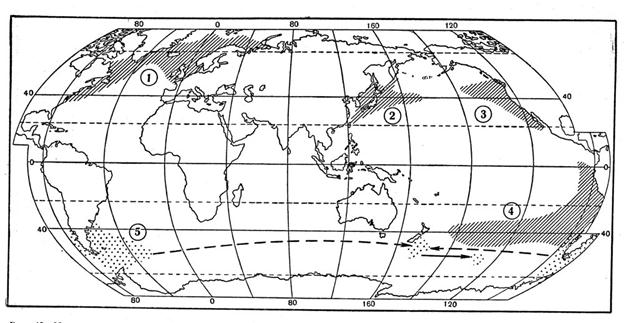 Рис, 15. Максимально расширенные ареалы псевдонеритических рыб — атлантической сельди (1), сардины иваси и дальневосточной скумбрии (2), калифорнийской (3) и перуано-чилийской ставриды (4) и возможные пути расселения южной путассу (3).
Рис, 15. Максимально расширенные ареалы псевдонеритических рыб — атлантической сельди (1), сардины иваси и дальневосточной скумбрии (2), калифорнийской (3) и перуано-чилийской ставриды (4) и возможные пути расселения южной путассу (3).
В двух последних случаях вид может рассматриваться как временно или постоянно обосновавшийся в том или ином конкретном участке открытого океана. Примеры, иллюстрирующие сказанное, ограничиваются, к сожалению, данными за самые последние десятилетия, так как имеющиеся ряды наблюдений очень коротки. Большая часть этих примеров относится к Северной Пацифике.
Известно, что в период высокой численности японского анчоуса и японской скумбрии в 50-е годы оба вида были весьма обычными и в открытых водах северо-западной части Тихого океана, в которых предполагалось даже существование их особых «стад» — в действительности, по всей вероятности, псевдопопуляций. В этом же районе в период высокой численности сардины иваси (Sardinops sagax melanosticta) в 30-е годы ее ареал расширялся вплоть до южной части Берингова моря (наблюдения в открытом океане отсутствовали), а современный подъем численности, начавшийся в 70-е годы (за первые 5 лет роста запас возрос не менее чем вдвое), привел к расширению области распространения в восточном направлении вплоть до центральной части океана, где, судя по некоторым сведениям, также образовывались временные псевдопопуляции. У японской скумбрии, как показано В. А. Беляевым, усиление воспроизводства на нерестилищах в прибрежной зоне Японии в середине 70-х годов привело к заселению океанических участков и формированию в районе Северо-Западного подводного хребта (гора Кинмей и прилегающие воды) зависимой популяции псевдонеритического типа: в 1978—1981 гг. скумбрия наблюдалась там на всех фазах жизненного цикла (икра, личинки, мальки, сеголетки).
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Т. symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955г. в удалении до 1100 миль, а в 1972 г.— до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставри-ды (Т. s. murphyi) в 40-х широтах южного полушария отмечен в 1985 г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45—60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2—3 лет). Таким образом, ати факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций), которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300—400 тыс. т в 1974—1976 гг. до 1100—1300 тыс. т в 1978—1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.