На примере аборигенов Австралии и Океании 3 страница
В пределах каждого крупномасштабного круговорота набор водных масс по вертикали специфичен, а это означает, что в каждом круговороте имеется своя структура вод. Принято различать поверхностную, подповерхностную, промежуточную, глубинную и придонную водные массы, которые разделены слоями воды с повышенными градиентами свойств (в пределах каждой массы свойства не меняются или меняются очень постепенно). Выделяют два главных типа гидрологических структур — тропические, расположенные в тропиках и субтропиках, и субполярные, расположенные в умеренных широтах.
Для тропических структур характерны следующие водные массы и слои раздела между ними. Сверху находится поверхностный однородный (изотермический) слой, который характеризуется высокой температурой (20—30°), мало меняющейся по глубине. Толщина этого слоя в среднем равна 50—100 м (в центральных частях субтропических круговоротов до 200—250 м). Под изотермическим слоем температура воды сравнительно резко надает, и ее градиент до глубины 400—500 м имеет порядок 0,1 град/м. Одновременно резко возрастает плотность воды. Под слоем повышенного градиента температура продолжает понижаться, хотя и более медленно. До глубины около 1000—1500 м (в зависимости от структуры) это понижение происходит, однако, быстрее, чем на еще больших глубинах. Этот слой называют «главным термоклином», и его нижняя граница выражена не очень четко. Глубже него вода отличается низкой температурой (около 2°), мало меняющейся с глубиной. Это так называемые «глубинные однородные воды».
Резкое расслоение толщи воды по температуре, а следовательно, и но плотности чрезвычайно характерно для тропиков. Плотность воды зависит, однако, и от солености, а в большей части тропической зоны выражен промежуточный слой повышенной солености на глубине нескольких сот метров, т. е. в верхней части главного термоклина. В результате главный термоклин распадается на два слоя слой максимума солености до глубин 200—400 м и слой менаду ним и границей глубинных однородных вод, обозначаемый обычно как промежуточный минимум солености.
В субполярных водах весь столб воды сравнительно однороден по температуре. Главного термоклина там практически нет, и набор водных масс существенно отличается от характерного для тропической зоны. Поверхностный изотермический слой может нагреваться летом выше 10° (при толщине около 50 м) и охлаждаться зимой до 0—5° (иногда и до отрицательных значений температуры). Под этим слоем летом может существовать промежуточный холодный слой, в котором температура удерживается на уровне ее минимального зимнего значения; он может достигать глубины около 300 м. Промежуточный теплый слой с температурой на 1—2° выше, чем в предыдущем, располагается примерно до глубины 2000 м. Глубинные однородные воды мало отличаются по своим характеристикам от глубинных вод в тропических структурах.
Рассматривая вертикальную термическую структуру Мирового океана, нередко придают особое значение противопоставлению теплого поверхностного и холодного глубинного объемов водной толщи, называемых соответственно «термосферой» и «психросферой». Автор этих терминов — датский океанолог и ихтиолог А. Бруун принимал за границу таких слоев изотермическую поверхность 10°. Таким образом, в высоких широтах термосфера, естественно, отсутствует, в умеренных широтах она имеет сезонный характер, а в тропической зоне постояпна, но варьирует по толщине от 100 м в восточных районах океанов, характеризующихся подъемом глубинных вод, до 700 м в западных.
Каждый участок дна омывается водами того или иного слоя водной толщи, и поэтому дно по вертикали может подразделяться на те же участки, что и пелагиаль (тонкий слой воды у самого дна изучен недостаточно, и о его характеристиках судят в основном по характеристикам основного объема соответствующей водной массы). На дне океана выделяются, кроме того, участки (зоны), различающиеся режимом и скоростью осадконакопления, размерами частиц грунта и содержанием в нем органических веществ.
Почти все органическое вещество в океане первично продуцируется в приповерхностной зоне — в освещенных (эвфотических) слоях, где возможен фотосинтез, а затем разными путями проникает вглубь и служит той пищей, которая обеспечивает существование населения в лишенных света слоях воды и на дне. Роль органического вещества, создаваемого в лишенных света глубинах океана за счет первичных хемосинтетических процессов, невелика: оно составляет лишь доли процента от синтезируемого растениями.
Продукция фитопланктона определяется целым рядом факторов — интенсивностью солнечной радиации, содержанием биогенных солей (фосфатов, нитратов, силикатов) , толщиной верхнего однородного слоя, прозрачностью воды и т. д. Главным лимитирующим показателем практически везде служит все же концентрация в эвфотической зоне биогенных солей, основной запас которых находится в более глубоких слоях воды. Распределение этих солей в открытом океане обусловлено в первую очередь динамическими факторами, вызывающими подъем глубинных вод в верхний слой. В соответствии с этим районы наибольшей первичной продукции расположены в умеренных широтах, где обогащение поверхностных слоев питательными веществами происходит в результате зимнего конвективного перемешивания, а также в некоторых районах тропической области, характеризующихся наиболее интенсивным подъемом «свежих» глубинных вод. К ним относятся районы поднятия вод (апвеллингов) в зонах действия сгонных ветров у берегов, дивергенции течений (особенно субэкваториальная дивергенция) и центральные участки некоторых локальных круговоротов. Наименьшими показателями продукции фитопланктона отличаются обширные по площади области опускания вод в антициклонических субтропических круговоротах. Ежегодная суммарная величина первичной продукции составляет, по современным представлениям, не менее 60 млрд т углерода.
Продукция фитопланктона, представляющего собой самый низший трофический уровень в пищевых взаимосвязях населения пелагиали, служит основой общей биологической продуктивности океана и в конечном счета его промысловой продуктивности. Дальнейшее перераспределение органического вещества, синтезированного водорослями-продуцентами, осуществляется животными консументами — растительноядным и хищным зоопланктоном, а затем нектоном — и проходит, таким образом, через несколько трофических уровней. При переходе на каждый последующий уровень расходуется большая часть энергии (такие потери часто достигают 90%), и это ограничивает число уровней: в холодных водах их 3—5, в тропиках — до 7. В одних пелагических сообществах (их называют сбалансированными по трофическим циклам) продукция, ежегодно создаваемая на каждом уровне, нацело выедается хищниками следующего уровня, в других (несбалансированных) эта продукция длительно и существенно превышает выедание и ее избыток выносится течениями в другие участки или попадает на дно. Наименьшей сбалансированностью отличаются неритические сообщества, а максимальное приближение к сбалансированности имеет место в тропической части открытого океана, где наблюдается высокая стабильность в количественном соотношении организмов, стоящих на разных трофических уровнях.
Количественное распределение и продукция зоопланктона в океане в общих чертах соответствуют обилию растительной жизни. Наибольшая биомасса среднеразмерного зоопланктона («сетного» зоопланктона, или мезопланктона) наблюдается летом в умеренных и высоких широтах (прежде всего во фронтальных зонах), а также в экваториальных районах и районах глобальных подъемов вод в восточных частях океанов, наименьшая биомасса — в зонах антициклонических субтропических круговоротов, так называемых «океанических пустынях». Наибольшая продукция мезопланктона также создается в субполярных водах обоих полушарий и в водах прибрежных и экваториальных апвеллингов, а иногда и в районах локальных продуктивных пятен.
Распределение биомассы океанического макропланктона и нектона тоже подчинено широтной и циркумконтинентальной зональности. Прослеживаются три полосы повышенной биомассы — в северных умеренных, экваториальных и южных умеренных широтах, причем в экваториальной зоне количественные показатели обилия снимаются в направлении с востока на запад в Тихом и Дтлантическом океанах и с северо-запада на восток — в Индийском океане. Эти полосы перемежаются четырьмя полосами пониженной биомассы — арктической, двумя центральными (субтропическими) и антарктической, в которой биомасса все же значительно выше, чем в трех других. Во всех океанах значительная биомасса консументов высших трофических уровней (главным образом рыб) приурочена к нерито-океанической зоне, переходной между прибрежными и океанскими районами. В этих участках максимальная биомасса нектона наблюдается на западных окраинах океанов в зоне полярных фронтов и на их восточной периферии мористее районов пассатных апвеллингов и вдоль прилегающих фронтальных разделов. В Индийском океане, где северного полярного фронта и северо-восточного апвеллинга нет, повышенная биомасса наблюдается мористее районов Аравийского и Сомалийского сезонных апвеллингов. В экваториальных широтах количество макропланктона и нектона особенно велико на востоке океанов (продуктивные зоны Гвинейского и Панамского заливов и прилегающих вод) и в районах локальных апвеллингов Карибского моря — Мексиканского залива и австрало-азиатских морей.
Все перечисленные выше (а также некоторые другие) особенности открытого океана оказывают непосредственное влияние на пространственное распределение обитающих в его водах организмов, в том числе и рыб. Участки, различающиеся по своему населению, могут иметь границы, совпадающие с изобатами, изотермами, изогалинами, линиями равного содержания растворенного кислорода, первичной продукции, биомассы мезоплаиктона и т. д., однако чаще всего эти границы соответствуют разделам дискретных водных масс, на которые подразделяется водная толща. Поэтому многие исследователи считают, что деление океана на крупномасштабные биотопы — участки, имеющие однородное население, — в первую очередь определяется именно расчлененностью его объема (так называемая «гипотеза водных масс»).
Границы водных масс в придонных горизонтах неплохо совпадают с основными формами рельефа дна. В вертикальном плане океан подразделяется поэтому на следующие зоны: верхнюю (от поверхности до глубины 100— 200 м), среднюю (до глубины 1000—1500 м), нижнюю (до глубины 3000 м), глубинную (до глубины 6000 м) и сверхглубинную (рис. 1). В классификации участков океанической среды обитания наибольшее значение имеет их разделение на пелагические биотопы, с одной стороны, и донно-придонные — с другой. В пелагиали верхняя зона соответствует поверхностным и подповерхностным водам, средняя — промежуточным, нижняя — глубинным глубинная — придонным, а сверхглубинная — водам глубоководных желобов; на дне (в бентали) верхняя зона более или менее совпадает с шельфом; средпяя и нижняя — с материковым склоном, глубинная — с ложем океана, а сверхглубинная — с дном желобов. При этом нужно иметь в виду, что приведенные примерный глубины не могут рассматриваться как абсолютные показатели границ вертикальных зон: их положение сильна варьирует в разных районах Мирового океана.
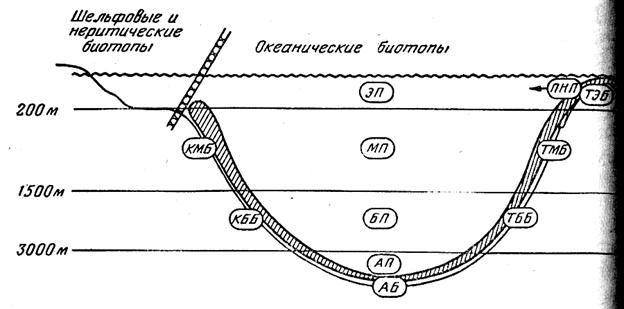 Рис. 1 . Вертикальная зональность океанических биотопо
Рис. 1 . Вертикальная зональность океанических биотопо
Названные вертикальные зоны в пелагиали вполне традиционно именуются эпи-, мезо-, бати-, абиссо- и хадопелагиалыо. Для бентали этих же зон в СССР обычна применяют названия сублитораль (на шельфе), «переходный горизонт» и батиаль (соответственно на верхней и нижней частях склона), абиссаль (на ложе океана) и ультраабиссаль (на дне глубоководных желобов). Им экбивалентны вертикальные зоны, названные Т. С. Рассом эпи-, мезо-, бати-, абиссо- и хадобенталью, и эта унифицированная терминология представляется более удобном для сопоставления биотопов дна и толщи воды (термин «батиаль» сохраняется при этом для обозначения всего биотопа материкового склона, т. е. идентичен понятию «мезобатибенталь»).
Население пелагиали контактирует с населением бентали во всех вертикальных зонах в придонных слоях водвой толщи, которые могут рассматриваться как биотоп вторичного сообщества — бентопелагического экотона (по определению К. В. Беклемишева, «экотон есть биотоп сообщества, состоящего из смеси видов двух соприкасающихся биоценозов»). В составе этого экотона в целом преобладают пелагические виды, а настоящие бентосные, как правило, находятся лишь па ранних стадиях жизненного цикла. В бентопелагиали имеются и специфические виды, и именно они составляют наиболее характерную часть ее населения. Это сообщество особенно четко обособлено в самой верхней вертикальной зоне (хорошо известное население неритической эпипелагиали), но оно существует во всем диапазоне океанских глубин. Бентопелагиаль, как и пелагиаль, является трехмерным биотопом и отличается в этом отношении от «плоской» двухмерной бентали, но объем придонной воды составляет ничтожную часть общего объема водной толщи, и в планетарном масштабе донный и придонный биотопы представляются тесно сопряженными (действительно, они не могут существовать в разделенном виде).
В дополнение к приведенной выше схеме вертикальной зональности А. П. Андрияшев выделил переуглубленные участки дна на шельфе — псевдобатиаль и изолированные от океанских глубин глубокие (более 3000 м) бассейны — псевдоабиссаль. Примеры псевдобатиального биотопа дают многочисленные депрессии на шельфе Антарктиды и в бассейне Северного Ледовитого океана, отделенные мелководьями от материкового склона; примерами псевдоабиссали (псевдоабиссобентали) являются Японское и Средиземное моря, соединяющиеся с океаном неглубокими проливами, обособленные котловины Полярного бассейна и т. д. Бесшельфовые подводные поднятия с глубинами порядка батиальных были выделены в талассобатиаль, и этот термин (также предложенный А. П. Андрияшевым) широко применяется в отечественной литературе. По аналогии с ним вершины подводных гор (часто их неправильно называют банками) с глубиной порядка 200 м и менее и «шельфы» малых островов были названы мной талассосублиторалью, или талассоэпибенталью. Кроме того, над высокими подводными поднятиями и около них (а иногда, по-видимому, и в других участках со слабо сбалансированными трофическими цикла ми) могут возникать условия, в некоторой степени ими тирующие особенности неритической зоны, и это позволяв ет обозначать такие участки как псевдонеритические (или талассоэпипелагические). Таким образом, в бентали и бентопелагиали Мирового океана три зоны — верхняя средняя и нижняя — существуют в двух модификациях субконтинентальной и талассной, а зона верхней пелагиали даже в трех — субконтинентальной (неритической) собственно океанической и талассной (псевдонеритической). Из семи субконтинентальных биотопов к океаническим относятся, однако, только донные и придонный биотопы материкового склона (соответствующие модифинации трех биотопов верхней зоны являются шельфо-нeритическими) (табл.1).
Эти наиболее крупномасштабные биотопы вертикального плана, как уже говорилось, делятся на биотопы подчиненных рангов, причем степень их возможного дробления в общем уменьшается от верхних слоев к нижним и от континентов к открытым водам. Из-за большего разнообразия условий в бентали степень дробности ее биотопов значительно больше, чем пелагических, которые как принято считать, представляют собой участки среды характеризующиеся наличием циркуляции, устойчивых в пространстве и времени.
Итак, система биотопических комплексов открытого океана имеет закономерную структуру и состоит из гомологичных частей. Примерами гомологичных биотопов в пелагиали могут служить парные крупномасштабным круговороты и их водные массы, расположенные симметрично по отношению к экватору в том же океане или занимающие сходное положение в других океанах (например, все субполярные или все центральные круговороты). В вертикальном расслоении водной толщи полная биотопическая гомология прослеживается только в пределах однотипных гидрологических структур — тропических и субполярных; так как если глубинные однородные по температуре воды (батипелагиаль) в обоих случаях вполне гомологичны, то поверхностный изотермический слоя в умеренных и высоких широтах в отличие от тропических имеет лишь сезонный характер. Поэтому нижняя граница эпипелагиали в общем совпадает с верхней границей скачка плотности воды (пикноклина), обусловленного в тропических водах понижением температуры, а в субполярных — увеличением солености. Мезопелагиаль в тропической зоне занимает слой основного термоклина — промежуточные воды с большими температурными градиентами, а за пределами тропиков — промежуточные холодные и теплые слои. Таким образом, тропическая и внетропическая мезопелагиаль сильно разнятся по условиям существования и не гомологичны между собой. Донные биотопы гомологичны между собой в той мере, в какой гомологичны водные массы, омывающие соответствующие участки дна. В пределах этих участков более дробные гомологи определяются типом осадконакопления.
| вертикальные зоны и их примерные границы по глубине | модификации биотопов | ||
| океаническая | талласная | субконтинентальная | |
| верхняя (от поверхности до 100-200м) | эпипелагиаль | эпипелагиаль, эпибенталь, эпибентопелагиаль | все биотопы заняты шельфо-неритическими сообществами |
| средняя (от 100-200 до 1000-2000 м) | мезопелагиаль | мезопелагиаль, мезобентопелагиаль | мезопелагиаль, мезобентопелагиаль |
| нижняя от 1000-1500 до 3000 м | батипелагиаль | батибенталь, батибетопелагиаль | батибенталь, батибетопелагиаль |
| глубинная(глубже 3000м) | абиссопелагиаль, абиссобенталь, абиссобентопелагиаль | не существует | не существует |
Сообщества, населяющие биотопы открытого океана, исключительно разнообразны в видовом отношении, но в этой книге они рассматриваются на примере только одного их фрагмента — таксоцена рыб, или ихтиоцена. Таксоценом называют совокупность всех видов некоторой таксономической группы любой надвидовой категория входящих в состав одного биоценоза любого иерархического ранга. Необходимость в таком термине определя ется тем обстоятельством, что изучение конкретных сообществ крайне редко проводится по полному состав слагающих его видов. В этом, действительно, не всегда есть нужда, так как многие особенности биоценозов могут быть исследованы и на примере отдельных таксоценов. В отечественную ихтиологическую литературу понитие «ихтиоцен» было впервые введено именно для обозначения совокупности рыб как части единого сообщества, объединяемого общим биотопом. Впоследствии в этом же биоценотическом смысле употреблялись такие термины, как «ихтиоценоз», «рыбная часть сообщества»; «рыбное сообщество», «рыбное население» и «ихтиокомплеке», из которых первые четыре должны рассматриваться как полные синонимы «ихтиоцена», а последний лишен строго детерминированного смысла. Некоторое представление о составе и разнообразии ихтиоценов открытого океана дает рис. 2.
Основные океанические ихтиоцены — пелагический на бентический (донный) — традиционно рассматриваются в качестве самостоятельных категорий. Этого нельзя сказать о бептопелагическом (придонно-наддонном) ихтиоцене, реальность существования которого над материковым склоном и ложем океана часто недооценивается, несмотря на его полную гомологию сообществу рыб неритической эпипелагиали.
Бентопелагические рыбы образуют довольно разнородную ассоциацию, в которой представлены виды разного происхождения и разной экологии. Среди них могут быть выделены 1) рыбы, обитающие в узком (толщиной в несколько метров) слое над дном, 2) рыбы, живущие в толще вод над дном, но не только вблизи дна, но и в зная чительном (на десятки — сотни метров) удалении 3) рыбы, мигрирующие ночью от придонных горизонтов в верхние слои водной толщи (эта группировка существует только в среднеглубинной зоне). Все бентопелагические рыбы почти не имеют прямого контакта с грунтом (некоторые, правда, откладывают донную икру или временами поедают донных животных) и этим отличаются от бентических рыб, которые могут лежать на дне, oпираться на него плавниками или ползать по нему, хотя и не теряют способности к плаванию. Для обозначения всей совокупности донно-придонно-наддонных рыб (при противопоставлении их чисто пелагическим видам) можно предположить название «бентопелагический комплекс».
По типу связи со средой обитания все океанические организмы группируются в три основных категории — планктон, нектон и бентос, причем рыбы представлены в каждой из этих жизненных форм. Планктон парит в толще воды и, не имея возможности длительно поддерживать направленную горизонтальную скорость, соизмеримую со скоростью перемещающихся вод, проявляет способность к активным миграциям только в вертикальном направлении. Нектон объединяет хорошо плавающих животных, «свободных в выборе своего пути» (по выражению Э. Геккеля) и произвольно совершающих не только вертикальные, по и горизонтальные перемещения, которые иногда достигают очень большой протяженности. Живущие на дне организмы — бентос — подразделяются па две группы: одну образуют сессильные формы (сидячие или малоподвижные), другую — вагильные, или мигрантные (способные к перемещениям по дну). Конечно, типичные представители всех трех основных категорий связаны многочисленными переходами: многие рыбы принадлежат, например, к нектопланктону и нектобентосу. Кроме того, многие животные бентоса и почти все нектонные животные проходят в своем развитии планктонную стадию. Плавучие икринки и личинки костистых рыб объединяются, например, под собирательным названием «ихтиопланктон».
Ихтиопланктон открытого океана включает не только ранние стадии развития океанических рыб (естественно, исключая щележаберных — акулы и скаты с самого начала жизни принадлежат к нектону), но и личинок и мальков множества шельфо-неритических видов, которые попадают в эпипелагиаль в результате пассивного выноса. В тропических водах его характеризует эфемерность существования — ведь развитие икры продолжается, как правило, не более 2 суток (правда, у макрелещуковых и летучих рыб — 2 недели, у вогмеровых рыб — до 2 месяцев), а личинки находятся в составе планктона в среднем около 8 недель. В более холодноводных районах эти сроки удлиняются, и длительность предмалькового периода жизни достигает у большинства видов 4—6 месяцев. Сильно растянуто личиночное развитие и у некоторых тепловодных форм, в частности у многих камбаловых и угреобразных рыб (личинки европейского угря, например, проводят в ихтиопланктоне до 3 лет), и это сильно способствует их дальнему разносу и широкому pacnpoстранению.
Икринки и личинки имеют приспособления к планктонному существованию, способствующие парению в толще воды: разного рода выросты и придатки на теле, удлиненные лучи плавников, а также жировые включения и газовые пузырьки (рисунок 3). Как правило, они сильно отличаются по всему своему облику от взрослых особей и в ряде групп претерпевают резкий метаморфоз (изменение строения) в ходе индивидуального развития. Поэтому установить принадлежность раннего малька, а тем более личинки (не говоря уже об икринке) к тому или иному виду, роду и даже семейству во многих случаях совсем непросто.
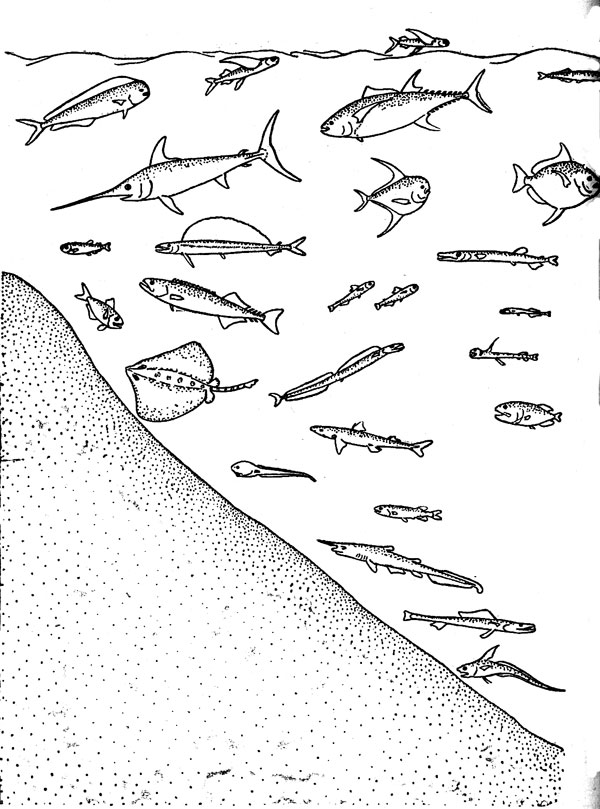
Рис. 2. Характерные рыбы океанических ихтиоценов (схема)

Рис. 2. Характерные рыбы океанических ихтиоценов (схема)
Особенно своеобразны прозрачные листовидные личинки угрей и близких к ним групп рыб. В прошлом столетии этих «рыбок» относили к особому роду Leptocephalus, и это название конечно, не в качестве родового осталось за ними до сих пор. Лептоцефалы некоторых видов достигают очень солидных размеров - рекорд (184 см) принадлежит L. giganteus (эта личинка относится к отряду спиношипообразных). Чтобы вырасти до такого размера, нужно, казалось бы, неплохо подкормиться, но, как ни странно, в кишечниках у этого гиганта или каких-либо других лептоцефалов никогда не находили даже следов пищи. Предполагают поэтому, что они могут усваивать растворенное в воде органическое вещество непосредственно через эпидермис. Согласно другому предположению, лептоцефалы высасывают протоплазматические соки из планктонных организмов, протыкая их покровы острыми зубами.

Рис. 3. Личинки океанических рыб
...В одном из первых тропических плаваний «Витязя» по судовой трансляции как-то прозвучало сенсационное объявление: «Сотрудниками ихтиологического отряда только что впервые в практике советских морских экспедиций выловлен экземпляр меч-рыбы. Сейчас рыба находится в лаборатории, где ее может увидеть каждый желающий». Через несколько минут помещение маленькой ихтиологической лаборатории было забито возбужденным народом, а толпа опоздавших теснилась у двери и иллюминаторов, выходящих на палубу. Они считали себя обманутыми, так как обещанной огромной рыбины, вооруженной мощным мечом, нигде не было видно. И тем не менее в объявлении не было неправды: в плоской чашке на столике бинокулярного микроскопа лежала полосатая игловидная личинка, усеянная мелкими шипиками и не превышавшая полутора сантиметров в длину... Через несколько лет при ином стечении жизненных обстоятельств эта моч-рыба не обманула бы ничьих ожиданий...
Мне никогда не приходилось всерьез заниматься изучением пхтиопланктопа (для этой исключительно тонкой работы нужны особые качества, которыми я не обладаю), хотя не раз доводилось иметь с ним дело, особенно в первых экспедициях — в те давно прошедшие времена одна из моих лаборантских обязанностей состояла в отборе икры и личинок рыб из планктонных проб. Эта нудная, как тогда казалось, работа (она и вправду очень однообразна) дала мне, однако, возможность воочию познакомиться с изумительным многообразием строения рыб на ранних стадиях развития, когда они так разительно непохожи на своих родителей. Я не могу забыть изумления и восхищения, которые мне доставляли тогда первые встречи с личинками листовидных совершенно прозрачных угрей, шарообразных колючих рыб-лун, стебельчато-глазых идиакантов, исключительно разнообразных по форме тела миктофид (взрослея, они становятся гораздо более схожими между собой), камбал, тунцов, рыб-топориков и прочими удивительными существами, которых так занимательно разглядывать под увеличением. С тех пор я сохраняю интерес к ихтиопланктону (мне даже удалось впервые определить видовую принадлежность нескольких неописанных личинок). Как и многие другие ихтиологи, я твердо убежден, что признаки ранних стадий рыб должны обязательно приниматься в расчет при изучении их систематики и филогении.
Итак, реальность существования и относительная обособленность планктона и нектона, несмотря на их не всегда четкое разграничение, не вызывают сомнений. Эти группировки представляют собой две крупные жизненные формы пелагического населения — пассивную и активную, из которых каждая по-своему хорошо приспособлена к обитанию в толще воды. Ю. Г. Алеев предложил использовать для разделения планктона и нектона безразмерный физический показатель — соотношение инерционных эффектов и вязкости, характеризующее процесс движения и называемое числом Рейнольдса (Re). Плавание планктона, по его мнению, происходит в режиме Re<2,0*10M>7 (как правило, Re<5,0*10>5), а нектона в режиме Re>5,0*103 (как правило, Re>105), причем первый случай как бы соответствует приспособлению к существованию в условиях ламинарных токов воды, а второй — в условиях турбулентных течений.
Форма тела нектонных рыб соответствует их активному образу жизни. Наиболее быстрым и подвижным хищникам — тунцам, акулам и др. — свойственна возникшая конвергентным путем торпедообразная форма тела. У некоторых рыб (меч-рыба, марлины) оно снабжено; спереди еще особым рострумом-обтекателем, который, как предполагает В. В. Овчинников, турбулизирует набегающий поток и, подобно выступу в подводной части носа современных быстроходных судов, сильно уменьшает лобовое сопротивление. Важную роль в обеспечении возможности быстрого движения имеют и хвостовые кили, увеличивающие поперечную жесткость хвостового стебля и выполняющие роль горизонтальных стабилизаторов. Для нектоирых рыб довольно обычно также симметрично сжатое с боков тело, в той или иной степени вытянутое в длину, или стреловидное тело с отставленными кзади спинным и анальным плавниками.
В открытом океане минимальные размеры нектонных рыб могут, по-видимому, находиться в пределах от 15 до 20—30см Еще раз повторю, что разграничение планктона и нектона остается очень условпым. Я думаю, что ни размеры, ни крейсерские скорости, ни числа Рейнольдса сами по себе недостаточны для разграничения этих жизненных форм. Нужно учитывать пространственный масштаб биотопа — нектон небольшого озера и нектон открытого океана не могут, по моему мнению, иметь одинаковые характеристики..
Планктонные рыбы обладают весьма ограниченными; возможностями для активного плавания на всех стадиях жизненного цикла. К этой группе принадлежат не только карликовые (длина 2—5 см) рыбки, населяющие глубокие слои пелагиали — циклотоны и другие представите ли семейства гоностомовых (Gonostomatidae), меламфайя мелкие светящиеся анчоусы и т. д., но и мпогие более крупные (длина до 30—50 см) малоактивные обитателя этого биотопа — нитехвостые угри, мешкороты, хаулиоды и др. К планктону (частично к нектопланктону) относятся и все рыбы, движущиеся при помощи ундулирующих, (колебательных) движений плавников — удильщики, рыбы-луны, сельдяные короли. А ведь некоторые из них; достигают очень внушительных размеров — нескольким метров. Такие рыбы почти не способны противостоять встречному движению воды, и это делает их объектом пассивного горизонтального переноса течениями.