Отступая от текста 10 страница
Самцы всех видов удильщиков отличаются от самок отсутствием иллициума, намного меньшими размерами, более прогонистым телом, сильным развитием обонятельных органов, наличием своеобразных вторичных зубов, сидящих на внешней стороне челюстей, направленных вперед и действующих как щипчики. В большинстве семейств они, несмотря на свою карликовость (длина 15—25 мм), ведут вполне активную самостоятельную жизнь, питаясь мелкими планктонными рачками и отыскивая самок по запаху. В то же время представители четырех семейств — Caulophrynidae, Ceratiidae, Linophrynidae и Neoceratiidae — демонстрируют совершенно уникальные отношения между полами, не известные ни у каких других позвоночных животных. Встретив самку своего вида, самцы этих удильщиков сначала накрепко вцепляются в нее зубами, а затем полностью срастаются с ее телом, теряя ненужные теперь органы (челюсти, глаза, кишечник), подключаясь к ее кровеносной системе и превращаясь в результате этих преобразований в самых настоящих паразитов, выполняющих только одну функцию — продуцирование спермы. Как правило, различия в размерах самок и паразитических самцов очень впечатляющи: так, на крупной церации длиной 119 см и массой 7 кг был обнаружен самец длиной 16 мм и массой всего 14 мг (по-видимому, он лишь недавно обрел свою «хозяйку», так как у других особей того же вида самцы-паразиты вырастают за счет самки до длины 16 см). У криптопсара известен, однако, случай прикрепления 10-миллиметрового самца к совсем юной, еще не половозрелой самочке длиной 15,5 мм, находившейся, по примерной оценке, в возрасте между 6 и 12 месяцами. Наибольший возраст крупной самки удильщика гигантактиса определен в 18 лет.
К макропланктону есть основания относить и саблезуба (Anoplogaster cornuta), отдаленно напоминающего удильщиков коротким телом, угольно-черной окраской и огромными клыками, но принадлежащего к отряду бериксообразных рыб. Мальки саблезуба по внешнему виду сильно отличаются от крупных особей: они имеют довольно светлую коричневатую окраску, почти треугольное в поперечном сечении туловище и «рогатую» голову, украшенную длинными заостренными шипами. Долгое время молодых и взрослых саблезубов относили к разным видам и даже родам, и лишь недавно было установлено, что индивидуальное развитие этой рыбы сопровождается столь существенными изменениями внешнего облика. Этот вид широко распространен в мезопелагиали тропической зоны, но размножается только в центральных круговоротах. Он достигает примерно 15 см в длину (при возрасте около 4 лет) и питается ракообразными, мелкой рыбой и кальмарами. Молодь, обитающую на глубине порядка 50—200 м, нередко поедают тунцы и другие хищные рыбы.
В мезо- и батипелагиали довольно обычны также представители семейства живоглотовых (Chiasmodontidae) — черные или коричневые бесчешуйные рыбы длиной до 30 см со слабой мускулатурой и огромным ртом, вооруженным многочисленными клыковидными зубами. Наиболее характерная особенность вошедшего во все учебники черного живоглота (Chiasmodon niger) — способность заглатывать крупную добычу, превышающую по размерам самого хищника (рис. 24). Челюсти, стенки тела и желудок у этого и близких видов могут сильно растягиваться. Светящиеся живоглоты (виды рода Pseudoscopelus), возможно, достойные обособления в отдельное семейство, отличаются наличием мелких фотофоров, расположенных отдельными группами на нижней поверхности тела — на горле, у брюшных и грудных плавников и у начала анального плавника. Живоглоты долгое время считались весьма редкими рыбами, и лишь сравнительно недавно было установлено, что виды этого семейства (в основном псевдоскопелы) играют важную роль в питании тех тунцов и марлинов, которые опускаются для откорма к границам основного термоклина. При изучении состава пищи большеглазого тунца в западной части Тихого океана, например, живоглоты были в том или ином количестве найдены больше, чем в половине всех исследованных желудков.

Рис. 24. Хиазмод, или черный живоглот, заглотивший крупную добычу
Еще одна группа пассивных макропланктоиных рыб, изучаемых сейчас Дж. Пэкстоном,— отряд китовидообразные, по общему абрису тела действительно несколько напоминающие миниатюрных настоящих китов. Они особенно характерны для батипелагиали, но редко попадают в руки исследователей; в мезопелагиали живут только ронделетии (род Rondeletia с 2 видами). Эти темно-красные, коричневые или черные рыбы, имеющие голое тело, большую голову с огромным ртом, маленькие или рудиментарные глаза и прекрасно развитую сейсмосенсорную систему, ведут, по всей вероятности, хищный образ жизни. Некоторые из них (например, виды рода Cetomimus), по-видимому, обладают способностью к люминесценции (светящимися органами считаются скопления губчатой ткани у анального отверстия и вдоль оснований плавников). Размеры большинства видов не превышают 10— 15 см, но самые крупные особи барбуризии (Barbourisia rufa) достигают 30—32 см в длину.
Переходное положение между макропланктонной и микронектонной группировками занимают эверманнелло-вые (семейство Evermannellidae) и жемчужноглазковые (Scopelarchidae) рыбы, недавно ревизованные Р. Джонсоном.
Эверманнеллиды — мезопелагические рыбы, длина которых варьирует от 7 до 18,5 см, с мощным зубным вооружением и довольно слабой туловищной мускулатурой. В состав семейства входят три рода, хорошо различающиеся строением глаз — у одонтостомопса (Odontostomops normalops) они имеют нормальное строение, у кок-корелл (род Coccorella) — полутелескошгческие, а у эверманнелл (Evermannella) — настоящие трубчатые, ориентированные вверх со слабым наклоном вперед. Обитающие в том же диапазоне глубин (300—1000 м, редко глубже) и тоже пользующиеся преимуществом бинокулярного зрения жемчужноглазки лмеют более упругое тело и, по-видимому, обладают лучшими возможностями для активного плавания. Длина самых мелких видов, ареалы которых приурочены к тропической зоне, составляет лишь 6—10 см (род Scopelarchus и др.), тогда как крупные представители рода Benthalbella, живущие в холодных водах Северной Пацифики и Антарктики, достигают 22— 24 см в длину. Характерной особенностью этих рыб, как и других, рассматриваемых ниже представителей подотряда алепизавровидных — веретенников, кинжалозубов, алепизавров,— является способ их размножения. Все они принадлежат к числу синхронных гермафродитов, гонады которых одновременно продуцируют как яйца, так и сперму. Таким образом, потенциально они способны к самооплодотворению, но никаких доказательств в пользу его реального существования нет, и при нересте партнеры скорее всего ведут себя так же, как и раздельнополые рыбы: один из них функционирует в качестве самца, другой — самки.
Семейство веретенниковых (Paralepididae) содержит, по А. Посту, 12 родов и около 50 видов, распространенных на глубине 200—1500 м (обычно 300—600 м) во всех широтах Мирового океана, но наиболее разнообразных в теплых водах. Особенно большой ареал, простирающийся от 40° с. ш. до 40° ю. ш. (отдельные нахождения и за пределами этой зоны, в Атлантике, например, вплоть до западной части Баренцева моря), у ложного нотолеписа (Notolepis rissoi), достигающего 30 см в длину. Другие виды семейства имеют длину от 15 см до 1 м (Sudis hyalina), и все они относятся к нектонной жизненной форме. Удлиненное тело веретенников прекрасно приспособлено для быстрого плавания и преследования добычи «в угон» (по этой причине они почти не попадают в тралы исследовательских судов), однако из подводных аппаратов наблюдали, как они медленно передвигаются в вертикальном положении головой вниз или вверх. Представители тропических родов Lestidium и Lestrolepis имеют светящиеся органы в виде одного или двух трубчатых каналов, расположенных в брюшной мускулатуре и, по-видимому, содержащих светящиеся бактерии. Веретенники питаются мелкими рыбами и различными беспозвоночными, но один вид (Notolepis coatsi), живущий в антарктических водах, специализирован на питании крилем. Сами они составляют очень существенную часть диеты тунцов и других крупных хищников.
Выделяемый в отдельное семейство, но близкий к веретенникам кинжалозуб (Anotopterus pharao) длиной до О см (этого размера он достигает в возрасте 6 лет) особенно характерен для умеренных вод северного и южного полушарий (крупные особи могут быть встречены даже у Гренландии и в Антарктике), хотя размножение идет, судя по ловам молоди, только в субтропической зоне. Кинжалозуб хорошо отличается от всех родственных ему рыб отсутствием спинного плавника, особенно резко контрастируя с алепизаврами (семейство Alepisauridae), обладающими огромным флаговидным плавником па спине.
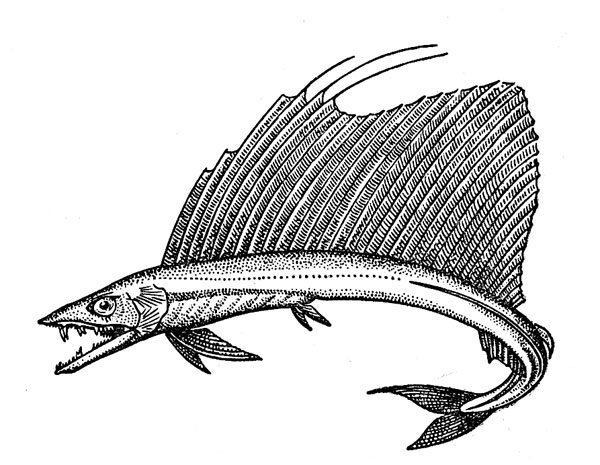
Рис. 25. Большой алепизавр.
Большой алепизавр (Alepisaurus ferox) (рис. 25) имеет очень широкое распространение. Он встречается как в тропических, так и в умеренных водах в очень растянутом диапазоне глубин — от подповерхностных горизонтов до 1000 м, будучи особенно обычным в слое от 150 до 350 м, и часто ловится на крючки тунцеловных ярусов. Максимальная зарегистрированная длина этой рыбы 210 см (второй вид рода — короткорылый алепизавр А. brevirostris не превышает 90 см). Экземпляры длиной 1,5 м и массой около 5 кг (по соотношению этих величин видно, насколько прогонисто их тело) вполне обычны в уловах. Алепизавры — прожорливые хищники, не очень разборчивые в выборе добычи. В списках их жертв значатся десятки видов рыб, головоногих моллюсков, ракообразных, а также сальпы, гребневики, медузы, пелагические полихеты и другие объекты. Следует отметить, что переваривание пищи у алепизавров (как и у родственных им рыб) происходит только в кишечнике, а желудок используется лишь в качестве своего рода «провизионной кладовой» и всегда содержит абсолютно целые объекты питания (из этого «орудия лова» происходит немало голотипов мезопелагических животных). Общее число жертв в желудке может быть очень велико: у одного алепизавра, например, там были найдены 41 рыба, 3 кальмара, 1 амфипода и 1 полихета, а у другого сразу 83 экземпляра рыб-топориков. Трофологические исследования показывают, что именно алепизавры являются главными пищевыми конкурентами «ярусных» тупцов, но они же принадлежат к числу первостепенных кормовых объектов этих тунцов.
Среди мезопелагических хищников нельзя не упомянуть небольших акул из преимущественно бентопелаги-ческого семейства катрановых (Squalidae), представляющих довольно специфический элемент океанической ихтиофауны. Их насчитывается около 10 видов. Акула-карлик (Euprotomicrus bispinatus) встречается во всех океанах и приурочена преимущественно к центральным водным массам (в других районах открытого океана ее замещает близкий и также карликовый вид — акула-пигмей Squaliolus laticaudus). Акула-карлик принадлежит к числу никтоэпипелагических рыб, мигрирующих ночью к поверхности (акула-пигмей не поднимается, как правило, выше 200 м), а в дневное время опускается, как предполагают, на глубину порядка 1000—1500 м. Самцы этого вида созревают при длине 17—19 см и достигают длины 22 см, самки немногим крупнее — самая большая из них имела размеры 27 см. Размножается путем яйцеживорождения, принося на свет 8 детенышей длиной около 6 см. Характерной особенностью акулы-карлика является способность к произвольному свечению. При возбуждении вся брюшная поверхность ее тела и нижние края боков светятся ровным бледно-зеленым светом, ярко вспыхивая при резких движениях рыбы и затухая, когда она успокаивается. Органами свечения служат мелкие фотофоры диаметром 0,3—0,8 мм, густо усеивающие всю нижнюю часть туловища. Люминесценция может быть очень интенсивной: известен случай, когда такую акулу заметили ночью на расстоянии 15 м от борта.
Близко к поверхности океана изредка встречается ночью и сигарная акула Isistius brasiliensis, широко распространенная в тропиках, но она, как правило, не поднимается выше верхней границы основного термоклина. Впечатляющий размах ее вертикальных миграций (с 3500 до 85 м) облегчается практически нейтральной плавучестью тела, обеспечиваемой большим количеством жидкого жира в полости тела и огромной печени. Эта акула, достигающая 50 см в длину (самцы несколько мельче — до 40 см), — прекрасный пловец. Наряду с потреблением в пищу относительно небольших рыб и головоногих она, как и второй вид рода — большезубая акула I. plutodus, способна к чрезвычайно своеобразному способу питания, который, по сути дела, может быть определен как факультативный паразитизм (уникальное явление в классе хрящевых рыб). Оба вида могут нападать на самых крупных рыб и китообразных. В этом случае акулы сначала присасываются толстыми губами к боку своей жертвы и вонзают в нее свои непропорционально большие нижнечелюстные зубы (рис. 26), имеющие форму треугольных лезвий и образующие в своей совокупности подобие острой пилы. Затем они производят вращательные движения, позволяющие выпилить из кожи и мышц жертвы конический или цилиндрический кусок, который извлекается из раны шиловидными верхнечелюстными зубами и препровождается в широкую глотку. Следы таких атак — характерные кратеровидные шрамы — отмечаются не так уж редко. Они были обнаружены на телах марлинов, тунцов, ваху, корифен, большеротой акулы, дельфинов и китов. Интересно, что такие же отметины были обнаружены при осмотре внешнего резинового корпуса американских атомных подводных лодок. Оба вида обладают способностью к люминесценции.

Рис. 26. Большезубая акула — факультативный паразит tspyniibix рыб и китообразных.
После лова в мезопелагиали сети и тралы иной раз приходят на борт с характерными дырами, происхождение которых долго оставалось непонятным — ведь в толще воды нет, как известно, предметов, за которые можно зацепиться. Оказалось, что эти разрезы - своеобразные автографы сигарных акул, побывавших в трале и благополучно ушедших на свободу. Лишь в крайне редких случаях, видимо, будучи пойманными в самых верхних горизонтах, они разрешают извлечь себя из воды, но и в этом случае чаще всего не ждут, пока их освободят из сети, а сами прорезают ее острыми, как бритвы, зубами, снова возвращаясь в море или, гораздо реже, падая па палубу.
Живые акулы (брать их в руки, как показывает мой опыт, надо с большой осторожностью), помещенные в аквариум, интенсивно светятся в темноте таинственным зеленоватым светом, и я видел эту иллюминацию несколько раз. Впервые такое свечение наблюдалось в начале прошлого века натуралистом старого закала Ф. Беппеттом, который оставил его прекрасное описание, приводимое здесь с некоторыми купюрами. «В темном помещении экземпляр представлял собой необычное зрелище, Вся нижняя поверхность тела и головы ярко излучала зеленоватый фосфоресцирующий свет, что поистине придавало животному страшный и пугающий облик. Люминесценция была постоянной и заметно не усиливалась при раздражении... Когда акула погибла (после трех часов наблюдений) люминесценция полностью исчезла... Моим первым впечатлением было, что рыба случайно получила некоторое количество светящегося вещества из моря, но более тщательное изучение не подтвердило этого подозрения, потому что постоянство, с которым свечение занимало определенные части тела и плавников, его неизменность при жизни и прекращение после смерти не оставляют сомнения в том, что это был жизненно необходимый процесс...»
Самыми крупными мезопелагическими хищниками являются, по-видимому, гемпиловые рыбы — змеиная макрель максимальной длиной около 1 м (как интерзональ-пый мигрант, она была рассмотрена в предыдущей главе), эсколяр (Lepidocybium flavobrunneum) длиной до 2 м (обычно 1—1,2 м), достигающая иногда еще больших размеров руветта (Ruvettus pretiosus). Оба последних вида более обычны, впрочем, над материковым склоном и подводными поднятиями и, может быть, должны считаться бенто-пелагическими. В своем вертикальном распространении они тяготеют к верхней части основного термоклина. Истинно мезопелагические темпилиды из родов Diplospinus, Paradiplospinus и Nealotus не превышают 25—45 см в длину.
В целом рыбы, входящие в состав глубоководных пелагических сообществ, характеризуются значительно меньшими размерами, чем рыбы приповерхностных слоев. Как показывают ориентировочные расчеты (в качестве примера проанализированы данные по максимальной длине приблизительно 400 видов, известных, согласно новейшей сводке, из вод Южной Африки), длина «среднестатического» глубоководного вида равна примерно 12 см, что на целый порядок меньше длипы «среднего» эпипелагического вида, которая достигает 102 см (рис. 27). При этом наибольшая длина двух третей (64%) мезобатипелагических видов не превышает 20 см, а преобладающей их части (85%) —50 см, тогда как абсолютное большинство (93%) эпипелагических видов может достигать Длины более 20 см, а почти три четверти (72%) — и более 50 см.
Неудивительно поэтому, что подавляющее большинство глубинных видов относится к пассивной жизненной форме (т. е. к планктону в широком понимании термина). К ней принадлежат и многие из крупных рыб (например, полутораметровый удильщик церация). Настоящие нек-тонные виды характерны только для самых верхних горизонтов мезопелагиали, а в холодных глубинах с ничтожными кормовыми ресурсами отсутствуют даже микронектонные рыбы: по-видимому, активный поиск и преследование добычи энергетически не оправдывают себя в этих специфических условиях.

Рис. 27. Распределение видов эпипелагических (J), глубоководных пелагических (3) и глубоководных донно-придонных (3) рыб по максимальной длине особей. Среднегеометрическая длина вида составляет 102 см в эпипелагиали, 12 см в глубинной толще воды и 37 см у дна и над ним.
Рассматриваемые в этой главе глубоководные пелагические рыбы входят в состав сообществ четырех крупномасштабных биотопов — мезо-, бати-, абиссо- и хадопелагиали, однако комплексы видов, населяющие эти биотопы, далеко не равнозначны по объему и разнообразию (табл. 3). Наиболее богатый видами (около 950) мезопелагический ихтиоцен достаточно хорошо обособлен от батипелагического (в нем около 200 видов), но с дальнейшим увеличением глубины происходит лишь общее обеднение фауны. Специфических абиссопелагических и тем более хадопелагических рыб, насколько можно судить по имеющимся сейчас данным, не существует, и поэтому ихтиоцены трех нижних зон водной толщи можно рассматривать совместно. Общее число видов в глубоководной, по-видимому, близко к 1000 (все цифры в табл. 3 следует рассматривать впрочем, как приблизительные).
Мезопелагический биотоп занимает, как уже указывалось, воды основного пикноклина, как правило, совпадающего с основным термоклином. Положение и четкость его верхней границы определяются в первую очередь температурным фактором. Именно поэтому в высоких широтах, где в зимне-весеннее время температура поверхностных и промежуточных вод становится практически одинаковой, нередко наблюдаются выходы глубоководных рыб даже к самой поверхности. В тропиках граница между мезо-и эпипелагиалью полностью проницаема только для никтоэпипелагических мигрантов. Эта граница, или, точнее говоря, пограничная зона, приуроченная к наиболее градиентным по температуре слоям и имеющая толщину порядка 100—150 м, должна рассматриваться как экотон — зона смешения видов из обоих контактирующих сообществ. Одни из этих видов проводят в нем определенную часть жизненного цикла, другие встречаются там постоянно или почти постоянно.
Для эпимезопелагического экотона особенно характерны крупные нектопланктонные и нектонные рыбы, находящие в этой зоне оптимальные условия существования. Некоторые из них — алепизавры, ряд видов веретенников и гемпилид — более тяготеют к верхней мезопелагиали, другие — упоминавшиеся в предыдущей главе сельдяные короли и их родичи, морские лещи, центролофы — к нижней эпипелагиали, но это разделение представляется весьма условным. Здесь же нагуливается во взрослом состоянии большинство крупных тунцов, мечерылых и акул, служащих объектами ярусного рыболовства. Основной источник пищи всех таких хищников — макропланктонные организмы (рыбы, кальмары, ракообразные), мигрирующие к верхней границе термоклипа ночью, однако те хищники, которые питаются преимущественно на утренних и вечерних зорях (алепизавры, тунцы и др.), могут использовать этот источник пищи только опосредованно — через постоянно живущих в мезопелагиали менее крупных хищных нектеров (это паралепидиды, гемпилпды и хиазмодонтиды из рыб и, возможно, некоторые кальмары).
Преобладающие в мезопелагиали макропланктонные и микронектонпые рыбы делятся па две основные экологические группы. Одну нз них (большую) образуют виды, совершающие регулярные суточные вертикальные миграции с ночным подъемом и дневным опусканием, другую — виды, сохраняющие во взрослом состоянии более или менее постоянный горизонт обитания.
Следует отметить, однако, что большинству представителей обеих групп (а также рыбам бати- и абиссопелагиа-ли) свойственны онтогенетические миграции: их икринки, личинки и мальки проходят развитие в слоях воды, расположенных ближе к поверхности, чем горизонты обитания подрастающих и взрослых особей. Проявлением одномоментного состояния онтогенетической миграции многих видов служит прямая связь размера рыб с глубиной обитания — в пределах вертикального диапазона, характерного для данного вида, более крупные особи распределены на большей глубине, чем мелкие. По мнению М. Е. Виноградова, основной смысл онтогенетических перемещений, которые хорошо выражены и у многих беспозвоночных, состоит в том, что растущие личинки и молодь могут откармливаться в более кормных верхних слоях воды.
Суточные миграции рыб (как и других пелагических животных) в самой общей форме обычно объясняют с трофических позиций, указывая, что вертикальные мигранты, поднимаясь в ночное время на меньшие глубины (вплоть до поверхности океана), используют в качестве пищи относительно более богатые ресурсы мезоплапктона этой зоны или поедают животных, перемещающихся вместе с ними. Такое объяснение ничего не дает, впрочем, для понимания обратного процесса — перемещения мигрантов на глубину после откорма. Для обоснования этого опускания было предложено несколько гипотез.
Так, в 50-е годы советский ихтиолог Б. П. Маитейфель, который был в пашей стране одним из основоположников изучения поведения рыб, высказал предположение, что сельди (а также другие рыбы из числа зрительных планктофагов) активно уходят вниз, спасаясь от «пресса хищников» поверхностного слоя, и придавал этому уходу прямое адаптивное значение. Такое объяснение не может быть принято: как ни трудно количественно оценить упомянутый «пресс», он выражен в мезопелагиали никак не меньше, чем в эпипелагиали (это особенно очевидно в тепловодной зоне, где наблюдаются самые масштабные вертикальные миграции миктофид и других макропланктонных рыб). Другая теория, также исходящая из адаптивности миграций, предполагает активное опускание вниз с целью уменьшения энергетических затрат мигрантов в том случае, если они питаются в более теплой воде, чем та, в которой происходит переваривание их пищи. Эта гипотеза, приложимая в отличие от предыдущей только к тропическим водам, не проверялась балансовыми расчетами и вряд ли выдержит такую проверку в подавляющем большинстве случаев.
По мнению Ю. А. Рудякова, ритмические вертикальные перемещения пелагических животных могут быть следствием изменения их двигательной активности в течение суток, причем опускание на глубину происходит в те часы, когда уровень этой активности оказывается недостаточным для компенсации пассивного погружения под действием силы тяжести. Для вертикально мигрирующих рыб наиболее правдоподобно выглядит следующая схема суточных перемещений: активный подъем вечером с целью встречи с главными кормовыми объектами, ночной откорм в слое обитания этих объектов (иногда с перерывом в полуночные часы), более или менее пассивное утреннее опускание на глубину (возможно, соответствующую переходу мигрантов в состояние нейтральной плавучести) и дневной пассивный отдых («сон») на этой глубине. В связи с этим нужно отметить, что большая часть мигрирующих рыб имеет плавательный пузырь — орган, довольно эффективно, хотя и не быстро регулирующий изменение удельной массы тела. У многих рыб вертикальные миграции начинают формироваться еще на личиночной стадии развития. Известно, например, что личинки многих массовых миктофид встречаются у поверхности воды только днем (это пример так называемой инвертированной, или обращенной, миграции), что прямо связывается с существованием у них суточного хода двигательной активности, обусловленной ритмом питания.
Общепризнано, что миграции контролируются суточными изменениями освещенности. Этот фактор может действовать как ориентирующий, сигнальный, определяющий ритм питания или синхронизирующий эндогенные (циркадные) ритмы отдельных особей.
Размах вертикальных перемещений может сильно варьировать. Наибольшей протяженности — с глубины 400— 800 (1000) м до поверхности — они достигают у никтоэпипелагических рыб, уже упоминавшихся выше. К ним относятся около 40 видов светящихся анчоусов из 5 родов, акула-карлик, змеиная макрель, которые, по-видимому, поднимаются к поверхности каждую ночь, а также отдельные виды винцигуерий и астронестов (например, Aslronesthes aiger), у которых такал миграция не имеет массового характера, и значительная часть популяции остается ночью в подповерхностных слоях,
У преобладающего большинства тропических мезопе-лагических рыб вертикальные миграции полностью укладываются в пределы своего биотопа — основного термоклина (т. е. между нижней границей поверхностного изотермического слоя и верхней границей глубинных однородных вод). При этом положение верхней границы ночного подъема разных видов довольно сильно варьирует: по-видимому, оно контролируется в основном температурой воды (может быть, также положением слоя концентрации пищевых объектов). Известную роль в размахе вертикальной миграции играет и освещенность (многие миктофиды, например, поднимаются в новолуние на 30— 50 м выше, чем в полнолуние). Имеются различия и в батиметрическом распределении разных размерно-возрастных групп одного вида, У некоторых видов (из родов Diaphus, Ceratoscopelus и др.) в нижние слои эпипелагиали проникают, как правило, только неполовозрелые особи.
Мигрирующие по вертикали рыбы перемещаются в толще воды в многовидовых ассоциациях — звукорассеивающих слоях, образующих особые сообщества неполного состава (без автотрофного звена), которые можно называть схизоценами. Эти слои объединяют рыб, креветок, эвфаузиид, кальмаров, киленогих и крылоногих моллюсков, сифонофор, сальп и других животных. Суточные миграции ЗРС (как и другие виды вертикальных перемещений) связывают мезопелагических консументов с пищевыми ресурсами вышележащих горизонтов и имеют существенное значение в перераспределении органического вещества в океане. Эти слои имеют очень широкое распространение и наблюдаются почти повсеместно. В продуктивных районах часто формируются сразу несколько звукорассеивающих слоев, перемещающихся по-разному, хотя иногда и сливающихся вместе ночью или днем. Основную роль в мигрирующих ЗРС играют макроплаыктонные рыбы-планктофаги, среди которых резко преобладают миктофиды из родов Diaphus, Ceratoscopelus, Lampanyctus, Hygophum и др., составляющие до 80—90% общей численности рыб в схизоцене. В некоторых участках океана к доминирующим формам относятся также винцигуерии и батилагиды. Хищные рыбы представлены в сообществах ЗРС хаулиодами, стомиями, гоностомами, астронестами, идиакантами и другими относительно крупными видами отряда Stomiiformes, питающимися только в пределах самого слоя. Глубины дневного и ночного положения звуко-рассеивающих слоев колеблются в довольно значительных пределах — у светящихся анчоусов, например, амплитуда суточных перемещений варьирует от 200 м (Diaphus regani) до 700—800 м (Ceratoscopelus warming!),— но почти всегда остаются в границах мезопелагиали, т. е. зоны, занятой промежуточными водами. В светлое время суток мигрирующие рыбы живут на глубине 300—1000 м (обычно глубже 400—500 м), а ночью концентрируются в слое от 50-100 до 300 м.
В мезопелагиали, как уже отмечалось, имеются и слабо мигрирующие рыбы, у которых размах вертикальных перемещений не превышает 50—100 м, т. е. настолько ничтожен, что им можно пренебречь (у некоторых, например у топориков рода Argyropelecus, ночью поднимается только верхняя граница). Есть там и рыбы, вовсе не мигрирующие на протяжении суток. Глубина обитания этих постоянно живущих на одном горизонте видов (их еще называют «стационарными») также не одинакова. Грубо говоря, на средних глубинах можно выделить два комплекса видов, разделение которых особенно четко выражено в тропических водах с развитым главным термоклином. Там, на глубине 400—500 м, температурный градиент сильно уменьшается и понижение температуры становится гораздо более медленным, чем в самом слое скачка, и это приводит к обособлению (конечно, не абсолютному) двух среднеглубинных группировок — верхне- и нижнемезопе-лагической. К первой относятся, в частности, рыбы-топорики рода Argyropelecus, эверманнеллиды, жемчужноглаз-ки, светящиеся живоглоты и др. Многие из них, можно повторить еще раз, входят в число основных кормовых объектов тунцов рода Thunnus и других крупных хищников. В нижней мезопелагиали постоянно живут «светлые» Циклотоны (например, Cyclothone alba), представители ро-Дов Melamphaes, Valenciennellus, Sternoptyx и др., и там же сосредоточиваются днем все рыбы, предпринимающие вертикальные миграции. На глубине порядка 300—400 м находится и нижняя граница ночного подъема наиболее слабо мигрирующих ЗРС. В результате верхние горизонты мезопелагиали — выше 300—500 м — в светлое время имеют очень скудное макропланктонное население, а макро-планктонные рыбы (если не считать личиночно-мальковых стадий развития) в этом слое днем отсутствуют.