Билет №8
1.Рибосоманың құрылысы
Рибосома (лат. rіbes — ағыс және грек. some — дене) — ақуыз синтезін жүзеге асыратын жасуша-ішілік органоид. Рибосомалар - екі орташа: үлкен және кіші өлшем бірлігінен тұрады. Нәруыздар биосинтезін (нәруызда аминқышқылдарды қосады) іске асырады.Рибосома тірі организмдердің барлығынан дерлік табылған. Әр жасушада шамамен 1000-нан 10000-ға дейін Рибосомалар болады. Шамамен диам. 20 нм-дей. Рибосомалар екі класқа бөлінеді: 70S және 80S (S-седиментация коэфф., Сведберг бірлігі). 70S Рибосома ядросы жоқ прокариоттар жасушаларында, ал 80S Р-лар эукариоттардың цитоплазмасында болады. Химиялық құрамы жағынан Р-лар РНІ-нан және ақуыздардан тұратын нуклеопротеид болып саналады. 70SBold text Рибосомасының 60 — 65%-ы РНҚ және 35 — 40%-ы ақуыздан, ал 80S Рибосомасының 50%-ы РНҚ-дан, 50%-ы ақуыздан құралады. Құрылымы жағынан Рибосомалар үлкен және кіші суббөліктерден тұрады.Рибосомалар цитоплазмада топтасып (5-70-тен) орналасып, полисомалар (полирибосома) түзеді. Рибонуклеопротеидтен құралған ( пен РНҚ) рибосомалар жасуша цитоплазмасында базофилді боялады, олардың құрамында ферменттер де кездеседі.
2.ДНК түзілуінің сипаттамасы
Организмде клеткалардың бөлінуі нәт-де өсіп-жетіледі. 1957 жылы М. Мезельсон мен Ф. Сталь, Дж. Уотсон және Ф. Криктің бактериялық клеткадағы ДНҚ-ның жартылай консервативті түрде екі еселенуі жөніндегі көзқарасын дәлелдеді. Сонымен, ДНҚ-ның еселенуі оның тізбектерінің ажырауынан басталады дедік. Ол тізбектерді геликаза (хеликс - спираль) - дезоксирибонуклеаза ферменттері - ДНҚ молекуласының бойымен екі бағытта жоғары және төмен ажыратады. Нуклеотидтер жұптарымен ДНҚ-ның шиыршықты тізбегінің арасындағы сутегінің байланыстары молекуланың бір жақ шетінде бірте-бірте үзіле бастайды және (ДНҚ) тізбектердің екеуі де бірінен бірі босай отырып, жазылады. Осылайша жазылған тізбек, өзінің қосылыстарын оське тік "қоя" отырып, дезоксирибоза және фосфор қышқылының қалдықтары арасында байланыстар арқылы ұсталып тұрады. Қоршаған ортадан клеткада жинақталған бос нуклеотидтер бар, олар ДНҚ-ның жазылған тізбегінің бос қосылыстарымен реакцияға түсе алады. Бірақ әр қосылысқа бір жұп, "толықтыра түсетін" нуклеотид қана жуықтап, жалғаса алады. Бұл жазылған тізбекке басқа, ДНҚ-ның жетіспейтін тізбегі жалғаса бастайды деген сөз. Осы процестердің нәтижесінде ДНҚ-ның екі молекуласы пайда болады. Олардың әрқайсысында қайтадан жинақталған молекуламен толықтырылған аналық молекуланың жартысы болады. Сонымен туынды молекулалар ДНҚ-ның аналық молекуласына мейлінше ұқсас келеді. Мұнда генетикалық материалдың құрамы да сақталады. Тізбектердің ажырауы мен қосылуы ферменттердің ықпалымен жүреді. Ажыраған тізбектерде оказаки фрагменттері жасала бастайды. Әр фрагмент он шақты нуклеотидтен тұратын РНҚ тізбегінен басталады. ДНҚ тізбегінің бойымен РНҚ түріндегі жаңа тізбекті праймаза (РНҚ - полимераза) ферменті ғана бастай алады. Тізбекті бастаған РНҚ бөлшегінен ары қарай "ДНҚ - полимераза-3" деген фермент ажыраған ДНҚ бөлігіне сәйкес етіп оказаки фрагменттерін синтездейді. Содан кейін басқа "ДНҚ - полимераза - 1" ферменті фрагменттердің бастаушысы болған әлгі РНҚ тізбегін ыдыратып жібереді. Енді кезек "ДНҚ - лигаза" деген ферментке келеді. Ол оказаки фрагменттерінің арасын ескі ажыраған тізбекке сәйкес етіп иуклеотидтермен толтырады. Ең соңында "ДНҚ полимераза-2" ферменті көптеген ферменттердің бірігуінен пайда болған жаңа тізбектің нуклеотидтерінің ескі тізбегімен сәйкес келетіндігін тексерді. Егер кандай да бір нуклеотид өз орнында тұрмаса соңғы аталған фермент оны кесіп алып тастап, оның орнына тиісті нуклеотидті қояды.
Осындай әр түрлі қызмет атқаратын ферменттердің үйлесімді жұмыс жасауы тұқымдық белгінің ДНҚ арқылы ұрпақтарға дұрыс өсірілуін қамтамасыз етеді. Әртрлі заттардың нәт-де клетка өңделгенде ДНК молек-да 2 есе көбейетіндігі дәлелденген.ДНК молекуласының екі еселеніп өсуі репликация д.а. Днк молекуласының репликациялану механизімінің нәт-де бір организмнің барлық клеткасының генетикалық мазмұны өзгеріссіз сақталады және гендер тұрақты болады. .
3.Транскрипция процесі,оның реттелу жолдары
ТРАНСКРИПЦИЯ (лат. transcrіptіo – қайта көшіріп жазу) – тірі жасушалардағы рибонуклеин қышқылының биосинтез процесі. Ол дезоксирибонуклеин қышқылы (ДНҚ) матрицасында жүреді. Транскрипция аденин, гуанин, тимин және цитозиннің қайталанбалы тізбегінен тұратын ДНҚ молекуласындағы генетикалық ақпараттың іске асуының бірінші кезеңі. Транскрипция арнайы ДНҚ және РНҚ полимераза ферменті арқылы жүреді. Транскрипция нәтижесінде РНҚ молекуласының полимерлі тізбегі түзіледі. Бұл тізбек ДНҚ молекуласының көшірілген бөлігіне комплементарлы болады.
Транскрипция процесінің өнімі әр түрлі қызметтер атқаратын РНҚ молекуласының төрт типінен тұрады:
1.рибосомадағы ақуыз синтезінде матрицаның рөлін атқаратын ақпараттық немесе матрицалық РНҚ;
2.рибосоманың құрылымдық бөлігін құрайтын рибосомалы РНҚ;
3.ақуыз синтезі кезінде генетикалық ақпараттың РНҚ-дағы нуклеотидтік “тілді” аминқышқылдық “тілге” ауыстыруға қатысатын тасымалдаушы РНҚ;
4.ДНҚ молекуласының репликациясы (генетикалық ақпаратты дәл көшіруді және оның ұрпақтан ұрпаққа берілуін қамтамасыз ететін нуклеин қышқылдары макромолекуласының өздігінен жаңғыру процесі) кезінде бастама қызметін атқаратын РНҚ.Транскрипция бірлігін атқаратын қызметі бір-біріне байланысты ферменттер синтезін анықтайтын гендер тобын ``оперон`` деп атайды. Прокариоттарда ол функционалды байланысқан бірнеше геннен, ал эукариоттарда тек бір ғана геннен тұрады. РНҚ-полимераза ферменті оперонның бастапқы бөлігін (промотор) “таниды”, онымен байланысып, ДНҚ молекуласының қос тізбегін ширатады. Осы жерден бастап мономерлі нуклеотидтер комплементарлы шартқа (принципке) сай РНҚ молекуласын түзеді. РНҚ-полимераза ферментінің ДНҚ-матрицасымен жылжуына байланысты синтезделген РНҚ молекуласы алшақтай береді де, ДНҚ-ның қос тізбегі қайта қалпына келеді. РНҚ-полимераза көшірілетін бөліктің соңына жеткенде (терминатор) РНҚ молекуласы матрицадан ажырайды. ДНҚ молекуласының әр түрлі бөліктеріндегі көшірмелер саны жасушаның қандайда болмасын ақуызды қажетсінуіне және қоршаған орта жағдайларына байланысты болады. Транскрипция процесінің реттелуін зерттеу молекулалық биологияның маңызды міндеттерінің бірі болып саналады. Ақпараттың көшірілуі ДНҚ молекуласынан РНҚ-ға ғана емес, сондай-ақ, кері бағытта, РНҚ-дан ДНҚ-ға да көшірілуі мүмкін. Мұндай кері Транскрипция құрамында РНҚ молекуласы бар ісік тудыратын вирустарда болады. Олардың құрамында жасуша зақымданғаннан кейін вирустың РНҚ-сын ДНҚ тізбегін синтездеуге матрица ретінде қолданатын фермент болады. Соның нәтижесінде ДНҚ молекуласының бір тізбегі, яғни ДНҚ – РНҚ гибриді түзіледі. Алғашқы РНҚ молекуласының барлық ақпаратын алып жүретін вируспен зақымданған қос спиральды ДНҚ молекуласы жасушаның хромосомасына еніп, қатерлі ісік тудырады. Кері Транскрипцияның ашылуы Ресей ғалымы Л.А. Зильбер (1894 – 1966) ұсынған қатерлі ісік вирусты-генетикалық теорияның дұрыс екенін дәлелдеді. Кері Транскрипция қалыпты жасушаларда ақпараттың жинақталуында және оның іске асуында (мысалы, эмбрионды даму кезеңінде) маңызды рөл атқаруы мүмкін
№ 9 емтихан билеті
1.Белоктың құрылымы мен функциясы жайлы бүгінгі таңдағы көзқарастар. Қазіргі кездегі түсінік бойынша белок дегеніміз – молекулалық массасы 5000 Д шамасынан көп, кеңістік құрылымы бар және организмде белгілі бір қызмет атқаратын полипептид. Белоктардың молекулалық массасы дальтон (Д) өлшемі бойынша беріледі. 1839 жылы голландия ғалымы Мульдер бұл заттың тіршіліктегі аса маңызына көңіл аударып белокты «протеин» (грек. Protos – ең басты, ең маңызды) деп атауды ұсынды. Белок химиясының тарихы, бәлкім 1745 ж. "Белок ғылым және өнер институтының комментарийларында (хабарла-маларында)" итальяндық Я. Беккаридің жарияланған жұмысымен басталған бо-лар. Бидай ұнынан бұрын белгісіз зат алғанын ғалым осы еңбегінде хабарлаған. Бидайдан алынған клейковина ұқсас болған.
Осылайша алғашқы белок препараты дайындалған. Сөйтіп, белоктық заттарды зерттеудің ұзында және өте қиын тарихы басталды. Ғалымдар ең қарапайым белок - инсулиннің молекуласының құрылысын ашқанша 200 жылдан артық уақыт өтеді. Осы мезгіл бойы белок құрамы, оның ұйымдасу жөнінде ондаған болжамдар ұсынылып және олар бекерге шығарылған. Барлық болжамдар ғылыми мұрағаттарда жайлы орын тапқан.
Әйтеуір, 1945 ж. көптеген зерттеушілердің күшімен белок молекулаларының бір ұзын тізбекке жалғанған амин кышқылдарынан тұратыны анықталды. Белок молекуласын түзетін амин қышқылдары көп емес - барлығы 20. Ал қараңыздар, белоктар молекуласы амин қышқылдарының кезектесіп орналасу бір ізділігімен ерекшеленеді. Мысалы, бір белокта валиннен кейін триптофан келеді. Олардың орнын ауыстыру арқылы басқа қасиеті бар жаңа молекула алуға болады. Айталық, егер біріншісі организмде маңызды қызмет атқарса, екіншісі мүлде қажетсіз болуы мүмкін.
2. Нуклеозидтер және нуклетидтер.Олардың атқаратын қызметтері.
Нуклеозидтер — азот негізі мен рибозадан (рибонуклеозидтер)немесе дезоксирибозадан (дезоксирибонуклеозидтер) тұратын қосылыстар. Олар пурин немесе пиримидин негіздерінің Ы-гликозидтері. Фосфокиназалардың әсерімен фосфорланып нуклеотидтер құрайды.
Азот негіздері рибоза, немесе дезоксирибозамен қосылып нуклеозид түзеді.
НУКЛЕОТИДТЕР – нуклеин қышқылдарын құрайтын негізгі құрылымдық бірлік. Құрамына азотты негіз, рибоза мен дезоксирибоза және фосфор қышқылының қалдығы кіретін қосылыс нуклеотид деп аталады.
Н. бір-бірімен ковалентті байланыс арқылы байланысқан 3 түрлі хим. бөліктен тұрады: 1) көміртектің бес атомы бар қант (ДНҚ молекуласында дезоксирибоза, ал РНҚ-да – рибоза); 2) қанттың 1-көміртек атомымен ковалентті байланысқан пуриндік немесе пиримидиндік азоттық негізі. Бұл екі бөлік бірігіп нуклеозид деп аталатын құрылым түзеді. ДНҚ молекуласының құрамына пуринді негіздер – аденин (А), гуанин (Г) және пиримидинді негіздер – цитозин (Ц), тимин (Т) кіреді. РНҚ-ны да осы негіздер құрайды, тек тиминнің орнына мұнда урацил (У) болады (Т урацилден метилдік топ [–СН3] арқылы ажыратылады). Н-дің 3-бөлігін фосфатты топтар (фосфор қышқылының бір немесе бірнеше қалдығы) құрайды. Фосфатты топтар бір қанттың 5¢-көміртегі атомымен келесі қанттың 3¢-көміртегі атомы арасында фосфодиэфирлік байланыстар құру арқылы полимерлі тізбек түзеді. Н-ді кейде нуклеозид-5¢-монофосфат деп атайды. Мыс., аденин негізді нуклеотидті аденозин-5¢-монофосфат, гуанин негізді нуклеотидті гуанозин-5¢-монофосфат, т.с.с. Н. түссіз кристалл, суда жақсы ериді, органик. еріткіштерде ерімейді, металдармен, негіздермен тұз түзеді. Н. жоғары энергиялы қосылыстар болғандықтан организмдегі энергия алмасу мен жинаудағы маңызы зор. Нуклеотидтер - ДНҚ мен РНҚ-ның құрам бөліктері. ДНҚ молекуласындагы әрбір нуклеотид үш компоненттен тұрады: тимин, аденин, гуанин, және цитозин, азот қосылыстары, 5 мүшелікөміртекті көмірсулар, дезоксирибоза және фосфор қышқылының қалдығы. РНҚ, да көмірсулардан рибоза бар, ап азот қосылысынан тимин урацилмен алмасқан.
3. Ферменттер әсер ету механизмдері. Олардың практикада қолданылуы.
Химиялық реакциялардың жылдамдығы реакцияға түсетін молекулалардың соқтығысу жиілігіне байланысты. Ал соқтығысу жиілігі молекулалардың концентрациясы мен ортаның температурасына байланысты. Температураның артуына қарай молекулалар қозғалысының кинетикалық энергиясы да артады, бұл молекулалар соқтығысуы-ның жиілігіне әсер етеді. Сонымен қатар реакдияның өтуі үшін молекулалардың соқтығысуы жеткіліксіз. Бұл кезде олар активті күйде болуы қажет, басқаша айтқанда, оларда реакция үшін қажетті энергияның біршама артық қоры болуы тиіс. Мұндай энергияны активация энергиясы деп атайды.Фермент осы реакцияға қажет активацияэнергиясын кемітеді. Ол үшін фермент реакцияға ұшырайтын заттың молекуласымен (оны субстрат деп атайды) бірігіп комплекс түзеді. Комплексті қысқаша Ф + С (фермент + + субстрат) деп белгілейді. Бұл комплекстің түзілуіне әлдеқайда аз мөлшердегі энергия қажет. Фермент + субстрат аралық комплексін түзу кезінде субстрат молекулалары біраз деформацияға ұшырайды, сондықтан реакцияның активация энергиясы кемиді. Бұл деформация субстраттың молекула ішілік байланыстарын әлсіретеді және молекуланы белгі-лі бір реакцияға неғұрлым қабілетті етіп шығарады. Комплекстің түзілуі спектрлік методтардың жәрдемімен дәлелденген.
Ф + С аралық комплексін түзуде субстрат ферменттің бүкіл молекуласымен емес, оның активтік орталықтар деп аталатын жеке-легең учаскелерімен қосылады. Ферменттің әрбір молекуласында 1—2 активтік орталық бар екендігі анықталып отыр.
Активтік орталықтық кеңістіктік құрылысы мен химиялық табиғаты белгілі бір субстратқа ғана сай келетіндей болып қалыптасқан. Бұл фермент басқа субстратқа катализатор бола алмайды. Осы ерекшелік ферменттердің талдаушылық қасиетін белгілейді.
Органикалық заттар мен ферменттердің структурасын зерттей келе,Э.Фишер фермент пен субстраттың кеңістіктік сәйкестігінің жақындығы женінде қорытынды шығарып, фермент субстратқа құлпының кілтіндей сәйкес келеді деп сипаттады.
Ферменттер молекулаларының құрылысына қарай екі топқа бөлінеді:
1) тек қана ақуыздардан тұратын бір компонентті ферменттер;
2) молекулаларының құрамына ақуыздан басқа активтік немесе простетикалық топ деп аталатын ақуызсыз текті заттар кіретін екі компонентті ферменттер.
Бір компонентті ферменттерде активтік орталықтың ролің амин қышқылдарының бүйірлік радикалдары атқарады. Ақуыз — фермент молекуласының II және III деңгейлік структурасы жасалған кезде, бүйірлік радикалдар өзара жақындасады да, активтік орталығын қүрады. Мысалы, панкреатикалық рибонуклеазаның активтік орталығына гистидин-16-ның, лизин- 41-дің және гистидин-119-дық қалдықтары кіреді. Активтік орталықтың осы компоненттері-нін. кеңістіктік жақындасуын күрделендіре түсетін амин қышқылдары да ферменттер үшін маңызды роль атқарады. Ақуыз — фермент молекуласының құрамынан басқа амин қышқылдарын ферменттің активтік қасиетіне нұқсан келтірмей де ажыратып алуға болады.
№ 10 емтихан билеті
1.Белоктың құрылымы және оларды анықтау әдістері.
Тірі организмдегі белок молекуласының әрқайсысының өзіне ғана тән белгілі бір кеңістіктік құрылымы болады. Ондай кеңістіктік құрылымды конформация деп атайды. Өзінің конформациясына сәйкес белоктар: глобулярлы белоктар және фибрилярлы белоктар болып екі топқа бөлінеді. Глобулярлы белоктардың құрылымы шағын ғана, олардың полипептидтік тізбектері сфера немесе эллипсоид сияқты тығыздала бүктелген болады. Белок глобуласының бетіне негізінен амин қышқылдарының полярлы тобы және зарядталған атомдары жинақталады. Олар сумен әрекеттеседі. Мұндай белоктарға ферменттер, қанның, сүттің белоктары (альбуминдер, глобулиндер және басқалары) жатады.Белок глобуласының ішкі бөлігі полярсыз ортаға жатады, ол гидрофобты және амин қышқылдарының полярсыз қалдықтарынан құралған. Гидрофобты ішкі ядроға су өтпейді, оның сумен байланысы жоқ.Фибриллярлы белоктар тұрақты келеді. Суда және сұйық тұз ерітінділерінде ерімейді. Белоктағы полипептидтік тізбектер белдік ось бойында өзара параллель орналасады, сөйтіп ұзын талшықтар – фибрильдер түзіледі. Мұндай белоктардың молекуласы созылған жіп сияқты және одан көп полипептидтік тізбектер тұрады. Белоктың бірінші реттік құрылымы дегеніміз – оның полипептидтік тізбегінде амин қышықлдарының белгілі бір ретпен жалғасып орналасуы. Бұл – табиғаттың аса қажетті құбылысы. Ол жағдай генетикалық негізде детерминденген және белок молекуласының басқа 3 құрылымының түзілуіндегі негізгісі болып табылады. Амин қышқылдарының тізбектегі кездесуі барлық белоктардың, биологиялық қасиеттері, әдетте ұқсас болып келеді.Екінші реттік құрылым.Белок молекуласындағы полипептидтік тізбек түзу емес, спираль тектес. Спиральдың бір айналымына 3,6 шамасындай амин қышқылдарының қалдығы келеді, спиральдың әр қадамы 0,54 нм. Белоктың осындай спираль тектес түрін оның екінші реттік құрылымы деп атайды.Үшінші реттік құрылым.Белоктың екінші реттік құрылымын түзетін спиральда түзу емес, ол да өз кезегінде бұралып иіріледі. Ондай бұралу кейде өте күрделі құрылым жасайды. Бір полипептидтік тізбектегі барлық атомдардың кеңістікте шап-шағын глобула жасап орналасуы белоктың үшінші реттік құрылымы деп аталады.Белок молекуласы атомдарының кеңістікте орналасуын, анықтайтын жалғыз-ақ әдіс – рентгенқұрылымдық анализ. Төртінші реттік құрылым.Екі немес одан да көп полипептидтік тізбектен тұратын белоктар олигомерлер деп, ал полипептидтік тізбектер протомерлер немесе суббөліктер деп аталады. Суббөліктердің кеңістікте өзара орналасуын белоктың төртінші реттік құрылымы деп атайды.Мұндай күрделі қызмет атқаратын белокқа қан гемоглобині жатады. Қан гемоглобині әрқайсысы миоглобин сияқты 4 суббөліктен құралған.
2. Ферменттердің қасиеттері.
Ферменттердің аса тұрақсыздығына байланысты әсері де көптеген факторларға тәуелді келеді.
Температураның әсері. Ферменттің жоғары активтігі 36-40ºС байқалады. Папаин бұған жатпайды. Оның активтігі 80º та да білінеді, ал каталазаға деген қолайлы температура 0 және 10ºС арасында жатыр. Температура 80-100ºС қа жеткенде фермент өзінің катализдік қабілетін жоғалтады (инактивацияланады) , денатурацияға ұшырайды. Инактивация реакцияның ұзақтығына және табиғатына байланысты.
Кебір ферменттер құрғақ күйінде 120-190ºС салқындыққа дейінгі температураға төзімді келеді. Температураны біртіндеп 37ºС қа дейін жоғарылатса, олардың активтігі қалпына келеді. Ферменттің бұл қасиетін малды қолдан ұрықтандыруға арналған ұрық сұйығын (сперма) сақтау үшін пайдаланады.
Ортаның рН ның әсері.әрбір ферменттің өте жоғары активтік көрсететін қолайлы рН аймағы бар. Мысалы, пепсин рН 1,5-2,5; трипсин рН 8-9; сілекей амилазасы рН 6,9-7; уреаза pH 7,2-8; болғанда ең жоғары активтік көрсетеді.
Талғампаздығы (іріктеушілігі). Әрбір фермент, құылысы жағынан ұқсас тек белгілі бір субстратқа, немесе заттар тоына ғана әсер етеді. Яғни, қайдай д бір басқа заттарға емес, тап сол берілген затқа әсе етуге бейімділігін айтды. Әрбір фермент тек белгілі бір реакцияны ғана катализдейді. Мәселен, уреаза ферменті бір ғана несеп нәрінің ыдырау реакциясын катализдейді. Сахараза тек сахарозаны ыдыратады.
Талғаушылықтың үш негізгі түрін айырады:
Топтық талғаушылық деп ферменттің белгілі бір химиялық байланыстарды үзе отырып заттар тобына әсер етуін айтады. Мысалы, пепсин, трипсин белоктардағы пептидтік байланыстарды гидролиздейді:
Жекелеген талғаушылық деп фрмент тек белгілі бір затқа ғана әсер етеді. Мәселен, каталаза сутек асқын оксидін суға жән оттегіне ыдыратады.
Құрылымдық талғаушылық деп ферменттің тек бір ғана кеңістік изомеріне әсер етіп, оның оптикалық антиподтарын катализдемеуін айтады. Мәселен, фермент моносахаридтердің Д қатарын, табиғи амин қышқылдарының a қатарын ғана ыдырата алады.Ферменттің тағаушылық қасиетінің биологиялық маңызы өте зор, өйткені ферменттер осындай қасиетінің арасында зат алмасу процесін реттеп отырады. Олар рекцияның жылдамдығын қай заттар, қандай бағытта өзгертетінін, өзгеру жолдары белгілейді.
3. Мутацияның нәтижесінде адам геномының өзгеруі,сипаттамасы.
Тұқым қуалаушылық сияқты өзгергіштік те барлық тірі организмдерге тән. Өзгергіштік дегеніміз — организмнің бойындағы түрлі белгілер мен қасиеттердің сыртқы орта факторларының әсерінен өзгеруі, соған байланысты ол жаңа белгі-қасиеттерге ие болады немесе өзінің кейбір белгі-қасиеттерін жоғалтады. Өзгергіштіктің екі түрі бар, олар: 1) фенотиптік немесе тұқым қуаламайтын өзгергіштік, бұған модификациялық өзгергіштік жатады; 2) генотиптік немесе тұқым қуалайтын өзгергіштік; бұған мутациялық және комбинативтік өзгергіштіктер жатады.
Мутациялық және комбинативтік өзгергіштік тек қана генотиптің өзгеруіне байланысты және ұрпаққаберіледі. Сондықтан оларды генотиптік немесе тұқым қуалайтын өзгергіштік деп атайды. Мутациялық өзгергіштік – дегеніміз организмнің генотипінің өзгеруіне байланысты болатын, яғни тұқым қуалайтын өзгергіштік. Дарвин тұқым қуалайтын өзгергіштікті табиғи және қолдан сұрыптаудың алғы шарты деп есептеді. Бірақ Дарвиннің тұсында тұқым қуалаушылық туралы тәжірибе жүзінде алынған деректер және оның ұрпаққа берілу заңдылықтары белгісіз елі. Сондықтан әр түрлі формадағы өзгергіштіктерді тұқым қуалау тұрғысынан айыру мүмкін болмады. Өзгергіштіктің түрлері туралы мәселе XIX ғасырдың аяғы мен XX ғасырдың бас кезінде ғана ғылыми тұрғыдан дәлелденді. «Мутация» деген терминді ең алғаш 1901 жылы Голландия ғалымы Г де Фриз өзінің «Мутациялық теория » деп аталатын еңбегінде қолданды.
Мутациялар әр түрлі бағытта жүзеге асады. Олардың көпшілігі организмнің тіршілік қабілетін кемітіп жібереді. Кейде өлімге душар етеді, оны летальді мутация деп атайды. Жалпы мутацияны эволюциялық процесстің алғашқы материалдық негізі деп есептеу керек. Бірақ жеке мутациялар түрді өзгерте алмайды.
Геномдық мутация – дегеніміз жасушадағы хромосомалар санының өзгеруіне байланысты организмнің белгілері мен қасиеттерінде пайда болатын өзгергіштікті айтады. Геном дегеніміз гапплидті хромососадағы гендердің жиынтығы.Енді осы геномдық мутациянын пайда болу жолын қарастырайық. Хромосома санының тұрақтылығын және оның ұрпақтан ұрпаққа берілуін қамтамасыз ететін жасушаның бөлуін механизмдері митоз мен мейоз екендігі белгілі. Бірақ кейбір жағдайда бұл механизмдер бұзылады да, хромосомалар жасушадағы екі полюске теңдей ажырамайды. Соның салдарынан хромосома саны өзгерген жасушалар пайда болады. Геномдық мутация тұтас гаплоидты жиынтықтың немесе хромосомалар санының көбеюіне немесе азаюына байланысты. Организмдерді хромосомалар саны гаплоидты жиынтыққа еселеніп көбеюіне қарай – полиплоидты, ал еселенбесе – анеуплоидты немесе гетероплоидты организмдер деп атайды.
№ 11 емтихан билеті
1.Биологиялық жүйелердегі сутектік, иондық, ковалентті, белоктық- гидрофобты байланыстардың ролі.
Сутектік байланыс оң зарядты полюстікке ие болған сутек атомы мен теріс полюсті, электртерістігі күшті, бөлінбеген электрон жұбы бар (көбіне F, О, N, кейде CI, S) атомдар арасында түзіледі. Сондықтан сутектік байланыстың механизмін жартылай электростатикалық, жартылай донорлы-акцепторлы деп қарастыруға болады. Сутектік байланысқа белоктардағы карбонил тобы мен амин тобындағы сутектің арасында түзілген байланыс мысал бола алады. Бұл полинуклеотидтер молекуласында іске асатын молекула ішіндік сутектік байланысқа жатады. Ал химияда көбіне молекула- аралық сутектік байланыстар кездеседі. Оны этил спиртінің өзінен немесе оны суға араластырғанда жылу бөле жүретін процестен байқауға болады. Кіші молекулалы су, спирт, альдегид тәрізді заттардың қалыпты жағдайда сұйық күйде болуы еутектік байланыстың әсерінен. Сонымен қатар аммиак, фторсутек сияқтыгаз күйіндегі заттардың оңай сұйылуы олардың молекуласының арасында түзілетін сутектік байланыстар арқылы іске асырылады. Сутектік байланыспен байланысқан заттар молекулалық кристалдық торға ие болады.
Иондық байланыс катиондар мен аниондардың арасында электро-статикалық тартылу күшінің нәтижесінде пайда болады. Химиялық әрекеттескен атомдар 8 электронды тұрақты октет қабатқа ие болып, катион мен анионға айналу үшін олар күшті металдар мен бейметалдарға жатуы тиіс. Иондық байланысқан қатты заттар иондық кристалдық торға ие болады. Сондықтан олар қатты, берік, қиын балқитын заттарға жатады. Иондық байланыс көбіне нағыз типтік металдардың оксидтері мен гидроксидтеріне және барлық тұздарға тән. Әдетте, бір молекуланың ішінде байланыстың әр түрлі типтері кездеседі. Мысалы, күшті негіздерде (КОН, Са(ОН)2, т.б. ) металл катионы мен гидроксотоп арасында иондық, ал оттек пен сутек арасында коваленттік полюсті байланыс түзіледі. Оттекті қышқылдардың тұздарында да (K2SО4, СаСО3, т.б.) металл катионы қышқыл қалдығының анионымен иондық байланыспен байланысса, оттек пен бейметалл (С, S) арасында коваленттік полюсті байланыс түзіледі. Жалпы алғанда, химиялық байланысты типтерге жіктеу шартты сипатқа ие. Өйткені олардың түпкі негізі бір. Мысалы, иондық байланысты коваленттік байланыстың шекті түрі деп қарау керек.
Коваленттік химиялық байланыс атомдардың арасында ортақ электрон бұлты түзіліп, ортақ электрон жұбы пайда болғанда шығады. Әрекеттесуге қатысқан элементатомдары тұрақты әрі тиімді 8 электронды конфигурацияға (s2p6) ие болады, тек сутек үшін 2 электрон(s2) жеткілікті. Коваленттік байланыс екі түрлі механизммен түзіледі: бірі — алмасу, екіншісі — донорлы-акцепторлы. Алмасу механизмі атомдардың жұптаспаған электрондарының бұлттары өзара бүркесіп ортақ жұп түзгенде байқалады. Мысалы, сутек молекуласында s — s орбитальдар бүркесіп, полюссіз коваленттік байланыс түзеді. Хлорсутек молекуласында s-және р- орбитальдардағы жалқы электрондардың бұлттары бүркесіп, полюсті коваленттік байланыс түзеді, өйткені ортақ бұлт электртерістігі үлкендеу хлор атомына қарай ығысады. Азот молекуласындағы үш ортақ жұп р-орбитальдардағы жалқы электронды бірдей бұлттардың бүркесуінен туатындықтан, полюссіз болып шығады Үш байланыстың біреуі (ядроны жалғастыратын сызық бойынша бүркесуі) σ-, ал екеуі 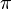 - (көлденең бұлттардың бүркесуі) байланыс болып саналады.Мұндай еселі байланыстар органикалық қосылыстарда жиі кездеседі. Қаныққан көмірсутек метанда 4 σ-байланыс бар, ал канықпаған көмірсутек этиленде бес σ-, бір -байланыс, ацетиленде үш σ-, екі -байланыс болатыны көрініп тұр.
- (көлденең бұлттардың бүркесуі) байланыс болып саналады.Мұндай еселі байланыстар органикалық қосылыстарда жиі кездеседі. Қаныққан көмірсутек метанда 4 σ-байланыс бар, ал канықпаған көмірсутек этиленде бес σ-, бір -байланыс, ацетиленде үш σ-, екі -байланыс болатыны көрініп тұр.
Донорлы-акцепторлы механизм бойынша коваленттік химиялық байланыс бір атомның дайын жұптасқан электроны екінші атомның бос орбиталіне орналасып, ортақ бұлттар түзгенде түзіледі. Ақуыз молекуласында аминқышқылдары бір-бірімен пептидтік байланыс арқылы байланысып үлкенді кішілі полипептид тізбегін түзеді.
Жоғары молекулалы қосылыс құрамына жүзге тарта пептидтік байланысы бар қосылыстар пептидтер деп аталады да, молекулалық массасы 10000 Дальтонды құрайды. Пептидтік байланысы жүзден асатын пептидтер ақуыздар деп аталады да, салыстырмалы молекулалық массасы 10000-нан бірнеше млн-ға дейін жетеді. Пептидтік байланысты аминқышқылдарының поликонденсациялануы деп қарастыруға да болады, яғни жоғары молекулалы қосылыс ақуызбен қатар, шағын молекулалы су түзіледі. Амиқышқылдарының қазіргі рационалды бөлінуі радикал полярлығына негізделген, яғни олардың сумен әрекеттесу қабілетіне байланысты. Ол амиқышқылыныңң төрт класын біріктіреді: 1) полярсыз-гидрофобты, 2) полярлы-гидрофильді-зарядталмаған 3)теріс зарядталған, 4)оң зарядталған. Келтірілген аминқышқылдар мыңдаған белоктардың құрамында әртүрлі мөлшер қатынасында болады, бірақ аталған аминқышқылдарының толық саны кейбір жеке белоктарда болмайды. Табиғи белоктардың көпшілігінде 20 аминқышқылы болады, кейбір белоктарда аминқышқыл туындылары: оксипролин, оксилизин, дийодтирозин, фосфосерин және фосфотреонин табылған. Бастапқы екі аминқышқылы біріктірілген ткань белоктарында кездеседі коллагенде ал дийодтирозин қалқанша безінің гормонының негізгі құрылысы болып табылады. Осы сияқты бұлшық ет миозинінде E-N –метилизин табылған.
2. Генетикалық акпараттың ДНҚ-да кодталуы.
Генетикалық ақпарат—организмдердің ұрпаққа беретін қасиеттері жөніндегі ақпарат.Генетикалық ақпарат нуклеин қышқылында оның негіздерінің кезегі түрінде жазылған. Ой жүзінде бұл әдіспен белокмолекуласының шексіз көп түрінің кодын жазуға болады. Генетикалық ақпарат бір ұрпақтан екінші ұрпаққа нуклеин қышқылының транскрипциясы арқылы беріледі. Генетикалық ақпарат өзгерісті, не өзгеріссіз түзетіліп сақталуы мүмкін. Бұған репарация, рестрикция, рекомбинация т. б. қатынасады.
Тіршіліктің барлық әралуандығын, клеткадағы әртүрлі биологиялық қызметтерді атқаратын түрлі ақуыз молекулалары қамтамасыз етеді.Ақуыздардың құрылысы пептидтік тізбектегі аминқышқылдарының жиынтығы және бір ізділікті орналасуымен анықталынады.Нақ осы аминқышқылдар бір ізділігі биологиялық немесе генетикалық кодтың көмегімен ДНҚ молекуласының тізбегінде шифрланған.
1954 жылы Г.Гамов, яғни ДНҚ молекуласындағы ақпараттың кодталуын бірнеше нуклеотидтер үйлесіп қамтамасыз етеді деп болжау айтты.
Өйткені, табиғатта әртүрлі 20 аминқышқылы бар, әралуандылықты ақуыздардан тұрады және оларды шифрлау үшін үш нуклеотид, немесе триплетті код қажет. Бұл жағдайда төрт нуклеотидтерден 43 = 64 триплет пайда болады. (Егер код екі нуклеотидтен тұратын болса, онда тек 16 аминқышқылын шифлаған болар еді, 42 = 16).
Генетикалық кодты толық шифрлау 20 ғасырдың 1960 жылдары іске асырылды. ДНҚ-ның 64 триплеті бар. Оның ішінде 61 триплет әртүрлі аминқыщқылдарын кодтайды, ал қалған үш триплет – мәнсіз немесе «нонсенс-триплеттер» деп аталады. Үшеуі аминқышқылдарын шифрламайды, олар ешбір аминқышқылына сәйкес келмейді (АТТ, АЦТ, АТЦ).
3.Мутация механизмдерінің реттелу жолдары.
Мутация (латын тілінде mutatіo – өзгеру) – табиғи жағдайда кенеттен болатын немесе қолдан жасалатын генетикалық материалдың өзгеруі. Соның нәтижесінде ағзаның белгілері мен қасиеттері тұқым қуалайтын өзгергіштікке ұшырайды. Ғылымға мутация терминін 1901 ж. голланд ғалымы Х. де Фриз (1848 – 1935) енгізді. Генетикалық аппараттың өзгеруіне байланысты мутацияның: геномдық, хромосомалық, гендік немесе нүктелік деген түрлері бар.
'Мутация (mutation) — жасушаның генетикалық материалының өзгеруі, бұл кейінгі ұрпаққа да беріледі.
Бұл тосыннан, кейде сыртқы факторлардың әсерінен болуы мүмкін (қараң. Мутагендер). Генетикалық кодты анықтайтын жүйедегі бір азоттық негіздің орнын басқа біреу алмастырса немесе бір немесе одан да көп негіздер генге енгенде немесе геннен жоғалғанда гендік мутация пайда болады. Мутациялардың көбі зиянсыз; оларды үнемі қалыпты доминанттық ген (қараң. Доминанттық) жауып тұрады.
Кейбір мутациялар айтарлықтай салдар туғызады; мысалы, ата-анасының екеуінен де тұқым қуалақшылықпен берілген белгілі бір мутация Орақ-жасушалы анемияныңпайда болуына әкеп соғады. Ұрпаққа жыныстық жасушаларда (аналық жасушасы немесе аталық ұрық) пайда болған мутациялар ғана беріледі. Әдетте, бұндай мутациялар ағзаға қауіпті.
Мутация ағза үшін тиімді өзгерістерге әкеп соғатын сирек кездесетін жағдайларда осы гені бар дербес ағзалардың саны мутацияға ұшыраған ген популяцияда қалпына келмейінше арта береді. Бұндай пайдалы мутациялар эволюцияның материалы болып табылады.
Геномдық мутация
Геномдық мутация – клеткадағы хромосомалар санының өзгеруі. Оның бірнеше түрі бар: 1) полиплоидия – хромосома жиынтығының бірнеше еселеніп өсуі; 2)анеуплоидия – хромосома жиынтығының еселенбей өсуі; 3) гаплоидия – диплоидты (екі еселенген) хромосома жиынтығының кемуі.
Хромосомалық мутация
Хромосомалық мутация – микроскоп арқылы көрінетін хромосома құрылымындағы өзгеріс. Бұл өзгеріс хромосоманың кей бөлімінің үзіліп қалуына (делеция), қосарланып кетуіне (дупликация) немесе оның басқа бір бөліміне ауысуына (транслокация) байланысты.
Гендік мутация
Гендік немесе нүктелік мутация деп ДНҚ молекуласының белгілі бір бөлігінде нуклеотидтердің қатар тізбегінің өзгеруін айтады. Ол молекулалық деңгейде өтеді, микроскоп арқылы көрінбейді. Мутация нәтижесінде ағза биохимиялық, физиологиялық, морфологиялық өзгерістерге ұшырайды. Организмдегі бұл өзгерістер бірден немесе біраз уақыттан кейін біртіндеп байқала бастайды. Полиплоидты мутанттардың клеткалары мен органдарының көлемі ұлғайып, хромосома жиынтығы жұп болса, оның ұрпақ беру қабілеті сақталады, ал тақ болса бұл қабілеті сақталмайды. Гендік мутация кезінде ағза үлкен өзгеріске ұшырайды. Кейде бір геннің өзгеруінен ағзаның бірнеше белгі-қасиеттері өзгереді (плейотропия). Гендік мутация доминантты (басыңқы),жартылай доминантты және рецессивті (басылыңқы) болады. Хромосомалық және гендік мутациялардың себебі көпке дейін белгісіз болып келді. Бұл өзгерістер ағзаға әр түрлі физикалық, химиялық факторлар – мутагендердің әсер етуінен пайда болады. Мысалы, радий сәулелерінің саңырауқұлақта тұқым қуалайтын өзгеріс тудыратындығын 1925 ж. орыс ғалымдары Г.А. Надсон (1867 – 1940) мен Г.С. Филлипов ашты. 1927 ж. АҚШ ғалымы Г.Меллер (1890 – 1967) жасанды мутацияның рентген сәулелерінің әсерінен болатынын тәжірибе жүзінде дәлелдеді. АҚШ генетигі С.Райт (1889 – 1988), орыс ғалымы С.С. Четвериков (1880 – 1959), ағылшын биологі Дж. Холдейн (1892 – 1964) қазіргі популяциялық генетиканың негізін салып, мутацияның эволюциялық мәнін ашты. Мутация көпшілік жағдайда ағза үшін зиянды болып келеді. Түрлі тұқым қуалайтын аурулар мен кемістіктерді тудырып, кейде тіпті өлімге душар етеді. Сонымен қатар кейбір мутациялар ағзаға пайдалы өзгерістер де алып келеді. Мысалы, гендік мутация (табиғи және қолдан сұрыптауға қажетті негізгі материал береді) өсімдіктер, жануарлар және микроазғаларды сұрыптау жолымен жаңа түрін алғанда кейбір қасиеттерін жақсартады.
№ 12 емтихан билеті
1.Нуклиен қышқылдарының химиялық құрамы..
Нуклеин қышқылдары (лат. nucleus — ядро) — құрамында фосфоры бар биополемерлер. Табиғатта өте көп тараған. Молекулалары нуклеотидтерден тұрады, бір нуклеотидтіқ 5'-фосфор арасындағы эфирлік байланысы мен келесі нуклеотидтің углевод қалдығының 3'-гидроксилі арасы эфир байланысымен нуклеин қышқылдары углеводты-фосфатты қаққасын калайды. Нуклеин қышқылдары жоғарғы полимерлі тізбектері ондаған немесе жүздеген нуклеотидтің қалдықтарынан тұрады. Олардың м. с. 105—1010. Нуклеин қышқылдары құрамына кіретін мономерлерінің (дезокси- немесе рибонуклеотидтер) түріне қарай ДНҚ жәңе РНҚ деп бөлінеді.[1]
Нуклеин қышқылдары тірі жасуша ядросының маңызды құрам бөлігі. Нуклеин қышқылдары (НҚ) рибонуклеин қышқылы (РНҚ) және дезоксирибонуклеин қышқылы (ДНҚ) болып екі үлкен түрге бөлінеді. Тірі организмнің құрамына нуклеин қышқылдарының екі түрі де кіреді. Нуклеин қышқылдары жоғары молекулалы гетерополимерлі қосылыстар.
уклеин қышқылдарының толық емес гидролизі нәтижесінде нуклеотидтер түзіледі (25-сызбанұсқа). Олар нуклеин қышқылдары полимер тізбегінде қайталанып отыратын күрделі құрылым буындары (монометрлері). Ал нуклеотидтерді одан әрі гидролиздесе, ортофосфор қышқылын және пентоза мен азотты негізге айырылатын нуклеозидтерді түзеді. Яғни, нуклеин қышқылдарының құрамына азотты негіздер (пиримидинді, пуринді), фосфор қышқылы және моносахаридтер (рибоза мен дезоксирибоза) кіреді. Нуклеин қышқылдары құрамындағы моносахаридтердің қалдығына байланысты рибонуклеин қышқылы және дезоксирибонуклеин қышқылы болып екіге бөлінеді. ДНҚ молекулалық массалары бірнеше мыңнан ондаған миллионға жетеді.
ДНҚ мен РНҚ құрамының айырмашылығы — нуклеин қышқылын толық гидролиздеу арқылы анықталды. Оларды гидролиздегенде, әр түрлі заттардың қоспасы түзіледі (36 кесте).
Нуклеин қышқылдары құрамында көмірсудың гидроксил тобы мен фосфор қышқылының арасында күрделі эфирлік байланыс түзіледі, ал азотты негіз көмірсудың жанынан жалғасады. Полинуклеотидтің құрылысын сызбанұсқамен былай өрнектеуге болады:
ақуыздар сияқты нуклеин қышқылдары әр түрлі болады. Олардың организмдегі функциясы да әр алуан. Нуклеин қышқылдарының да ақуыздар сияқты әр түрлі құрылымдары болады.
Нуклеин қышқылының бірінші құрылымында мононуклеотидтер белгілі тәртіппен орналасады.
Нуклеин қышқылының екінші құрылымы макромолекулалардың кеңістікте қос шиыршық болып орналасуын көрсетеді. Бұл кезде молекулалар арасында және молекула ішінде сутектік байланыс арқылы әрекеттесу болады.
НҚ-ның макромолекуласы екі полинуклеотидті тізбектен құралады. Олар кеңістікте қос оралма түзеді (54-сурет). Оралманы фосфор қышқылының полиэфирі түзеді, пиримидин және пурин туындыларының жазық молекуласы оралманың ішінде болады.
Нуклеин қышқылының макромолекуласындағы бірінің ішінде бірі жатқан ширатылған екі оралмада, пиримидин және пурин қалдықтары өзара сутектік байланыс арқылы байланыскан.
Сутектік байланыс белгілі бір жұп пиримидин және пурин туындыларының арасында түзіледі. Оларды комплементарлы жұптар деп атайды. Ондай жұптар: тимин (Т) — аденин (А) және цитозин (С) — гуанин (G).
ДНҚ-ның қос оралмалы сызбанұсқасында таспамен көрсетілгендері фосфор қышқылымен көмірсулардың полиэфирінің макромолекуласы. Бұларды қосып жатқан түзулер пиримидин және пурин туындылары, олар комплементарлы жұптар.
Нуклеин қышқылының үшіншілік щрылымы — ДНҚ мен РНҚ-ның кеңістікте шумақталып орналасуы.
2. Хромосома, оның кұрылысы және атқаратын қызметтері.
Хромосомалар (хромо… және грек. soma — дене) — жасуша ядросында болатын, гендерді тасымалдайтын және организмдер мен жасушалардың тұқым қуалау қасиеттерін анықтайтын органоидтар. Хромосомалар өздігінен көбейе алады, өзіндік атқаратын қызметі мен арнайы құрылымы бар және оны келесі ұрпақта сақтай алады. Хромосомалар терминін алғаш рет неміс ғалымы В.Вальдейер ашты (1888). Ол хромосомаларды негізгі бояғыштармен қарқынды боялатын тығыз денешік деп атады. Бірақ хромосомалардың сыртқы пішіні жасуша циклінің әр түрлі сатыларында өзгеріп отырады. Митоз және мейоз процестерінің метафаза кезеңінде хромосомалардың морфологиясы жарық микроскопының көмегімен анық көрінетін құрылымға енеді. Көптеген өсімдіктер мен жануарлардың дене жасушаларындағы хромосомалар ұрықтану процесі біреуін аналықтан, ал екіншісін аталықтан алған екі хроматидтерден (ұзынша жіпшелер) тұрады. Мұндай хромосомалар гомологты деп аталады. Мейоз процесінен өткен жыныс жасушаларында гомологты хромосомалардың тек біреуі ғана болады. Клеткадағы хромосомалардың толық жиынтығы кариотип деп аталады. Прокариоттар мен вирустарда хромосомалар болмайды. Оларда тұқым қуалау негізі ретінде әдетте бір жіпшелі немесе сақина тәрізді дезоксирибонуклеин қышқылы немесе рибонуклеин қышқылы болады және олар цитоплазмадан ядро қабықшасы арқылы оқшауланбайды. Клеткалық және тіршілік циклдері барысында хромосомалардың сыртқы көрінісінің өзгеріп отыруы олардың қызметінің ерекшеліктеріне байланысты. Ал хромосомалардың жалпы құ-рылымдық негіздері, биологиялық түрге байланысты әр түрлі болуы және ұрпаққа үздіксіз беріліп отыруы өзгеріске ұшырамайды. Бұған әр түрлі организмдердің хромосомаларын генетикалық, цитол. және биохимиялық зерттеулердің нәтижелері дәлел бола алады және олар тұқымқуалаушылықтың хромосомдық теориясының негізін құрайды. 1928 ж. хромосомалардың ең алғашқы молек. түрдегі үлгісін орыс ғалымы Н.К. Кольцов (1872 — 1940) ұсынды. Эукариоттардың хромосомалардағы ДНҚ молекуласы гистондық және гистондық емес белоктармен байланысып, кешен құрайды. Аталған белоктар ДНҚ-ның хромосомаларда жинақталып, оралған күйде болуын және жасушадағы РНҚ-ын синтездеу қабілетінің реттеліп отыруын қамтамасыз етеді (қара Транскрипция). хромосомаларға тұқым қуалау ақпаратының жазылуы ДНҚ молекуласының құрылымымен іске асырылады. Клеткадағы хромосомаларда ДНҚ молекуласының 99%-ға жуығы жинақталған, ал қалған 1%-ы басқа жасушалық органоидтарда (хлоропластар, митохондриялар) болады. Хромосомалар жасушада өте күрделі құрылымға ие және олар өте маңызды қызметтер атқарады. Хромосомалар құрылымын және қызметін зерттеу қазіргі заманғы биологияның өзекті мәселелерінің біріне жатады. Әсіресе, 20 ғасырдың 60 — 70-жылдары хромосомалар құрылымының молек. негізін түсінуге молекулалық генетиканың дамуына байланысты қол жетті. Бұл жаңалықтар тұқым қуалаушылықтың хромосомалық теориясындағы негізгі заңдылықтарды дәлелдеп, онан әрі дамытуға мүмкіндік береді.
3. ДНҚ полемераза ферментіне сипаттама.
Прокариоттық ДНҚ полимеразалар:
Ішек таяқшасы бактериасында және басқа прокариоттарда қызмет атқаратын ДНҚ полимеразалардың үш типі белгілі. ДНҚ полимеразалар 1 және полимераза 2 ДНҚ репарациясына қатынасады, ал ДНҚ полимераза 3 ДНҚ репликациясына қатынасады. ДНҚП күрделі фермент, оның 5 активті сайттары бар: 1. матрицалық сайт 2. праймерлік сайт 3. 5!-3! Ыдыратушы сайт немесе экзонуклеазалық сайт 4. Нуклеотит- үшфосфат сайт 5. 5!-3! Эндонуклеазалық сайт. ДНҚП 1 ферментінің атқаратын қызтеті синтездік активтілік көрсетіп, нуклеотиттерді ретімен қосып ДНҚ тізбегін өсірумен қатар экзонуклеозалық активтілігіне байланысты тізбектердегі қателіктерді жөндеп , түзету болып табылады. ДНҚП 2 активтілігі жағынан ДНҚ 1ге ұқсас болсп келеді, бірақ нуклеотиттерді қосып, ұзартып өсіру үшін тізбектің 3! Бос ұшына ғана әсер ете алады. ДНҚП ДНҚ репликациясында маңызды роль атқарады. Бұл мультимерлік фермент немесе холофермент суббірліктен тұрады.
Эукариоттық ДНҚ полимеразалар 1. ДНҚ полимераза альфа цитоплазмалық немесе үлкен полимераза деп те аталады, ол ядрода және цитолпазмада кездеседі. 2. ДНҚ полимераза бета нулеазалық полимераза кіші полимераза, тек омыртқылыларға тән фермент. 3. ДНҚ полимераза гамма митохондриялық полимераза, ядода синтезделеді. 4. ДНҚ полимераза дельта сүтқоректердің жасушаларынан табылған және PSNA белогіна тәуелді; ол ДНҚ синтезінің жылдамдығы мен қарқындылығын қамтамасыз етеді. 5. ДНҚ полимераза эпсилон қызметі бойынша дельта ДНҚ полимеразаға ұқсас, ашытқы саңырауқұлақтары мен сүтқоректілердің жасушаларында кездеседі ДНҚ молекуласындағы реаликация басталатын нүтеде А- Т жұптары көбірек, арнайы нуклеотиттер жүйесі орналасқан болады.
№ 13 емтихан билеті
1.Ферменттер, олардың биологиялық ролі, құрылымы және классификациясы.
клеткадағы биология лық ролі:
1.Ферменттер-клеткадағы химиялық реакцияларды жылдамдататын биология лық белсенді органикалық заттар. Кез келеген хим иялық реакциялардың жылдамдығы химиялық заттардың қасиетіне, олар дың концентрациясына, ортаның температурасына байланысты.
2.Клеткадағы химиялық реакциялардың ерекшелік тері – ферменттер деген биологиялық катализатор лардың қатысуымен өте жоғарыы жылдамдығы. 3.Ферменттердің химия лық табиғаты.Ақуыздар-ферменттердің міндетті құрамдас бөлігі. Барлық ферменттер-белоктар, бірақ белоктар фермент тер емес.
4.Ферменттің әсер ету механизмі. Химиялық реакциялардың ең басты шарты-реакцияға қатысатын молекулалар дың тығыз жақындасуы . Фермент құрылымының бұзылуы-оның катализдік қасиеттерінің жойылуы ның себебі.
Ферменттер бұл – күрделі органикалық молекулалар. Егер химиялық формуласын жазатын болсақ, мысалға, ұйқы безінің ферментін, оданда сандық мағынасы мынадай болады С1130Н1783О356N308S1
Басқа ақуыздар сияқты, ферменттер де біоіншілік, екіншілік, үшіншілік және төртіншілік құрылыммен сипатталады:
-біріншілік құрылымы – бұл пептидтік байланыстармен қосылған аминқышқылдардың реттк тізбегі;
-екіншілік құрылымы – полипептидтік тізбектің коныормациясын спираль түрінде СО- және NH- топтары арасындағы сутектік байланыстармен біріктіріп реттейді;
-үшіншілік құрылымы – бұл фермент млекуласының гидрофобтық ядроның түзілуімен полипептидті тізбектің глобулаға жиналуы. Оны химиялық байланыстардың тбарлық түрлерін тұрақтандырады. Коваленттік байланыстар – пептидтік және дисульфидтік; әлсіз – сутектік, гидрофобтық және электростатикалық.
-Төртіншілік құрылымы – бұл екі немесе бірнеше глобулярлық бірліктердің шоғырлануы (ассосация) нәтижесінде түзіледі.
Күрделі ферментте белоктық бөлікті (апофермент) және белокты емес компонентті (простетикалық топ, кофермент, кофактор) өледі. Ферменттің белоктық бөлігінде активті орталық – бұл субстратпен спецификалық байланысқан полярлы емес химиялық топтардың жиынтығы. Оның, полипептидтің басқа бөліктерінен айыратын топографиялық шекаралары болмайды. Активті орталықтарда, протондардың донрлы (-СООН, -NH3, -SH) немесе акйепторлары (СОО, -NН2, -S) болып табылатын ерекшеленген аминқышқылдардың қалдықтары орналасқан.
Классификациясы;
Қазіргі заманғы фермент тер классификациясы ар найы халықаралық Биохи миялық Одақ Комиссиясы мен әзірленген және “Но менклатура ферментов” атты кітапта жазылып, 1996 жылы орысша аудар масы шықты. Классифи кация негізінде үш ереже жатыр: а) катализденетін реакциялар түріне байла нысты барлық фермент тер 6 класқа бөлінеді.
1-класс – Оксидоредукта залар – тотығу – тотық сыздану реакцияларына жауапты
2–класс–Трансферазалар-химиялық топтардың бір субстраттан екіншісіне тасмалдауын катализдейді (мысалы, моносахариттер ді, амин қышқылдарын, фосфор қышқылдарын және т.б ). Бұл класқа протеинкиназа, ацетилтр ансфераза, фосфотрансфе раза және т.б жатады.
3–класс–Гидролазалар субстраттың химиялық қосылыстарын судың қатысуымен гидролиздей ді, ыдыратады. Бұл реак циялар теңдеуі:
RR1 + HOH → R – OH + R1 – H1; ( Амилазалар, протиназалар, лиазалар ) субстратқа химиялық топтардың ажыратуын немесе қосылуын катализ дейді ( аспартаза, фемира за және т.б).
4–класс–Лиазалар–2 бай ланыс түзу арқылы субс траттан белгілі бір топт тарды гидролитикалық емес ажырату реакциясын катализдейтін ферменттер немесе 2 байланыс үзілген жерден топтарды бірікті ру реакцияларын (мыса лы, H2O , NH3, және т.б) ажыратады.
5–класс–Изоимеразалар–субстраттың бір молекула сы ішінде құрылымдық өзгерістерді атқарады не месе изомерациялайды (глюкозоизомераза, три озофосфатимомераза жә не т.б )
6–класс–Лигазалар (синтетаза)-энергия жұм сап қарапайым қосылыс тардан күрделі қосылыс тарды синтездейді ( ДНҚ-геназа, аспарагин синтета за, пируваткиназа және т.б )б) әрбір фермент ке жүйелік атау беріледі, онда субстрат атауы, ка тализденетін реакция түрі, және “аза” жалғауы жал ғанады. Сондықтан фер менттер атауының екі ж үйесі қалыптасты.
в) әрбір ферментке төрт мәнді цифр беріледі. Бірін ші сан ферменттер клас ын, екіншісі класс тармағ ын, үшінші – клас тарма ғының тармақшасын, төр тіншісі – тармақшадағы ферменттің реттік нөме рін білдіреді.
2. Нуклеин қышкылдары, оның қасиеттері және құрылысы.
Нуклеин қышқылдарының толық емес гидролизі нәтижесінде нуклеотидтер түзіледі (25-сызбанұсқа). Олар нуклеин қышқылдары полимер тізбегінде қайталанып отыратын күрделі құрылым буындары (монометрлері). Ал нуклеотидтерді одан әрі гидролиздесе, ортофосфор қышқылын және пентоза мен азотты негізге айырылатын нуклеозидтерді түзеді.