Особенности регуляции синтеза белка у эукариот
Регуляция генной активности у прокариот
Схема регуляции активности генов на уровне транскрипции была впервые разработана Ф. Жакобом и Ж. Моно (1961) на примере лактозного (lac)-оперона кишечной палочки (E.coli). Единица регуляции транскрипции была названа опероном. Оперон – это последовательность структурных генов, определяющих синтез группы белков, участвующих в одной метаболической цепи, имеющих общий промотор и оператор.
E.coli быстро растет на питательной среде, содержащей в качестве источника углерода глюкозу. Глюкоза – самый доступный и выгодный для E.coli источник углерода и энергии. Пока глюкоза имеется в достаточном количестве, клетка не нуждается в получении и переработке других сахаров.
После переноса клеток на среду, содержащую вместо глюкозы лактозу, рост сначала замедляется, а затем возобновляется с высокой скоростью. При этом бактерии синтезируют три необходимых для усвоения лактозы фермента:
· β-галактозидазу, которая расщепляет лактозу на глюкозу и галактозу;
· пермеазу, необходимую для проникновения лактозы через клеточную мембрану;
· тиогалактозидазу.
Эти ферменты кодируются тремя структурными генами (соответственно Z, Y и А) лактозного оперона. Кроме структурных генов в состав оперона входят промотор и оператор. В схеме оперонной регуляции присутствует также ген-регулятор, кодирующий белок-репрессор. Однако ген-регулятор не входит в состав оперона и может находиться на расстоянии от него.
При отсутствии в среде лактозы активный белок-репрессор связывается с оператором, блокируя транскрипцию структурных генов. Хотя РНК-полимераза может связаться с промотором, она не перемещается дальше «выключенного» оператора (рис. 45А). При добавлении в среду лактозы ее молекулы связываются со специфическим участком репрессора, инактивируя его. У инактивированного репрессора резко снижается сродство к зоне оператора, в результате чего он отсоединяется, и РНК-полимераза начинает транскрипцию структурных генов (рис. 45Б).
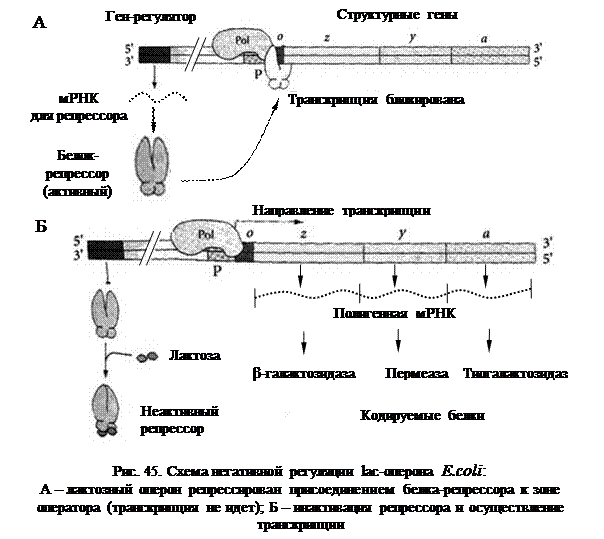
Снижение концентрации субстрата – лактозы – вновь является сигналом к соединению белка-репрессора с оператором и прекращению транскрипции генов lac-оперона. Таким образом, синтез ферментов происходит только в том случае, если в них возникает необходимость. Данная схема регуляции называется «негативной», так как контролирующим транскрипцию фактором является негативный фактор, «выключатель» – белок-репрессор. Индукция (включение) происходит при потере сродства белка-репрессора к оператору.
Наряду с негативной системой регуляции, для lac-оперона существует и позитивная регуляция. Промотор лактозного оперона способен связывать не только РНК-полимеразу, но и особый белок-активатор катаболизма (САР) в комплексе с циклическим АМФ (рис. 46А).
Присутствие САР и цАМФ вызывает не репрессию, а напротив, активирование транскрипции. Без САР РНК-полимераза не может связаться с промотором и начать транскрипцию. САР, образовав комплекс с цАМФ,
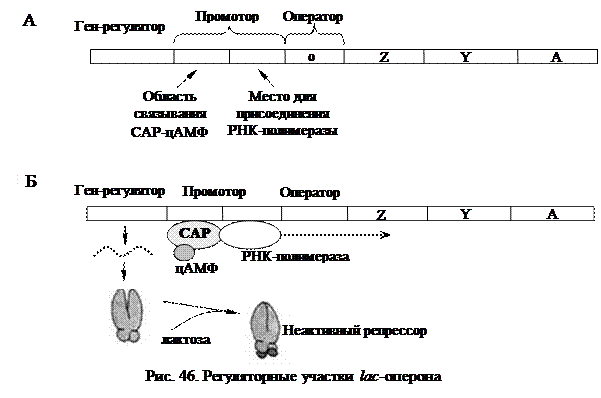
активизируется и только после этого присоединяется к своему сайту (САР-участку) на промоторе, многократно (почти в 50 раз) усиливая транскрипцию генов lac-оперона. При этом транскрипция возможна только в присутствии лактозы, когда оператор не блокирован репрессором (рис. 46Б). В случае присутствия в среде глюкозы, концентрация цАМФ в клетке резко снижается, и не образуется комплекса цАМФ с САР. В результате этого РНК-полимераза не может связаться с промотором и lac-гены не транскрибируются.
Механизмы регуляции биосинтеза белков у эукариот более разнообразны и осуществляются на разных уровнях. Так, стойкую репрессию генов вызывает компактная упаковка хроматина, включая взаимодействие с гистонами, образование нуклеосом и хроматиновых фибрилл. В гетерохроматине для транскрипции доступно менее 1% генов, в эухроматине, имеющем более рыхлую укладку, − значительно больше. В разных типах клеток в область эухроматина попадают неодинаковые гены, что обеспечивает стабильную репрессию одних генов и дерепрессию других на протяжении всей жизни клетки.
Большинство генов эукариот подвергается адаптивной регуляции, осуществляющейся на уровне транскрипции. Вследствие огромной протяженности и сложности эукариотической ДНК специфические регуляторные участки ДНК и взаимодействующие с ними белки-регуляторы весьма многочисленны. Выявлено более 100 различных белков, способных взаимодействовать с регуляторными последовательностями ДНК и тем самым влиять на сборку транскрипционного комплекса и скорость транскрипции. Эти белки содержат ДНК-связывающие домены, отвечавшие за узнавание специфических участков в молекуле ДНК, а также домены, активирующие транскрипцию. Последние связываются с транскрипционными факторами либо с РНК-полимеразой. Регуляторные белки могут иметь в своем составе антирепрессорные домены, которые взаимодействуют с гистонами нуклеосом, освобождая от них участки ДНК. Эти белки могут содержать в себе также домены, связывающие лиганды – индукторы транскрипции (стероидные гормоны, гормоны щитовидной железы, производные витаминов). После связывания лиганда конформация белка изменяется, и он образует участок, узнающий в регуляторной зоне ДНК специфическую последовательность и индуцирующий транскрипцию определенного гена.
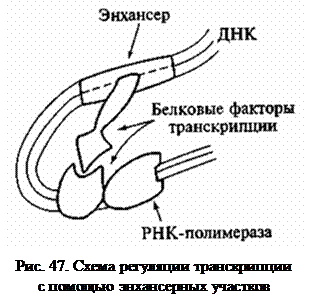 Важную роль в регуляции активности генов играют участки ДНК, расположенные на значительном (1000 и более пар оснований) расстоянии от промотора.
Важную роль в регуляции активности генов играют участки ДНК, расположенные на значительном (1000 и более пар оснований) расстоянии от промотора.
Энхансеры («усилители») – последовательности ДНК, служащие в качестве специфических участков связывания регуляторных белков, активизирующих процесс транскрипции.
Сайленсеры («глушители») – участки ДНК, которые, связываясь с белками, обеспечивают замедление транскрипции. Вероятно, влияние этих элементов на транскрипцию связано с изменением топологии цепей ДНК, в частности с образованием петель (рис. 47), что приближает регуляторные последовательности к промоторам, с которыми они взаимодействуют с помощью белковых факторов.
Регуляция на уровне транскрипции наиболее экономична, но она осуществляется недостаточно быстро. Поэтому важное значение имеет регуляция на других этапах реализации генетической информации.
На многих эукариотических генах, имеющих полиэкзонное строение, после транскрипции и процессинга образуется несколько вариантов зрелой мРНК, когда зкзон одного варианта сплайсинга может оказаться интроном в другом. Это приводит к образованию разных мРНК и, соответственно, разных белков с одного первичного транскрипта. Так, в парафолликулярных клетках щитовидной железы в ходе транскрипции гена гормона кальцитонина образуется первичный транскрипт мРНК, который имеет в своем составе шесть экзонов. мРНК кальцитонина образуется путем сплайсинга первых четырех зкзонов. Этот же первичный транскрипт в клетках головного мозга в ходе альтернативного сплайсинга образует другую мРНК, кодирующую белок, не обладающий гормональной активностью.
На состав белков клетки оказывает влияние также неодинаковая стабильность мРНК. Время жизни эукариотических мРНК составляет от нескольких часов до нескольких дней. Расположенный на 3'-конце фрагмент поли-(А) увеличивает продолжительность жизни молекул мРНК и, соответственно, количество белка.
Регуляция синтеза белка осуществляется и на уровне трансляции. Разные мРНК имеют неодинаковое сродство к рибосомным субчастицам, поэтому полирибосома может содержать различное количество рибосом. Так определяется соотношение белков в клетке. Наконец, может происходить подавление инициации трансляции всех мРНК клетки (например, при действии теплового шока, стрессах, недостатке железа, вирусной инфекции и т. п.). Стрессовый фактор индуцирует фосфорилирование второго фактора инициации (IF-2), тем самым инактивирует его и, следовательно, трансляцию.