Аксиома вторая
Все живые организмы должны быть единством фенотипа и программы для его построения (генотипа), передающегося по наследству из поколения в поколение.
Ничто не ново под луной. Еще в конце прошлого века биолог Август Вейсман сформулировал этот принцип (деление организма на сому и наследственную плазму).
Все последующие успехи генетики и теории информации лишь блестяще подтвердили его.
Многие читатели, возможно, пожмут, плечами: какая разница передается ли по наследству сама структура в виде маленького организма, запрятанного в яйцеклетке или спермии или же программа кодирующая его построение? Что дает новая теория развития (назовем ее генетической) по сравнению с преформизмом?
Что дает? Да все: мы сразу избавляемся от подавляющей картины бесконечной вереницы вложенных друг в друга зародышей. И не нужно отождествлять генетическую теорию развития с преформизмом, как это иногда делают (появился даже термин «неопреформизм»). Делая это, мы отождествляем программу построения структуры с самой структурой. Но это столь же нелепо, как отождествлять страницу из поваренной книги с обедом, рецепт – с лекарством и чертеж автомобиля – с самим автомобилем.
Как и в теории эпигенеза, упорядоченность организма в каждом новом поколении возникает заново.
Но упорядочивающий фактор – не мистическая энтелехия Аристотеля или «существенная сила» Вольфа. Это вполне реальная программа, закодированная, как мы теперь знаем, в длинных нитевидных молекулах дезоксирибонуклеиновой кислоты – ДНК или рибонуклеиновой кислоты – РНК у некоторых вирусов.

Рис. 6. Относительные размеры некоторых вирусов (электронная микрофотография с увеличением в 200000 раз). Слева направо – один из маленьких фагов φX174, вирус табачной мозаики, сложный бактериофаг Т4 (из группы так называемых Т‑четных фагов бактерии кишечной палочки).
Порядок организма возникает не из ничего, а из порядка полученной от родителей программы.
Подчеркнем одно обстоятельство. Наша трактовка первой аксиомы функциональна, она не связывает первое условие жизни с каким‑либо конкретным химическим веществом. Самовоспроизводящуюся машину можно в принципе построить, вкладывая в нее программу, записанную на магнитной ленте или в совокупности перфокарт или еще каким‑либо способом.
Принцип раздельного копирования при совместном существования генотипа и фенотипа остается незыблемым. То же и в жизни. В земных условиях основа фенотипа – белки, основа генотипа – нуклеиновые кислоты. Но не подлежит сомнению, что жизнь во Вселенной бесконечно разнообразна. Где‑нибудь на планете системы тау Кита или альфа Эридана жизнь может быть построена на иной структурной основе, но по единому для всей Вселенной принципу. Аксиома № 1 едина для всего живого. Жизнь на основе только одного фенотипа или же одного генотипа невозможна, при этом нельзя обеспечить ни самоподдержания, ни самовоспроизведения сложной специфической структуры.
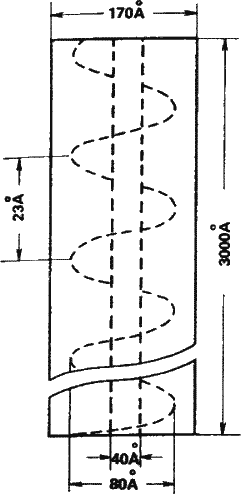

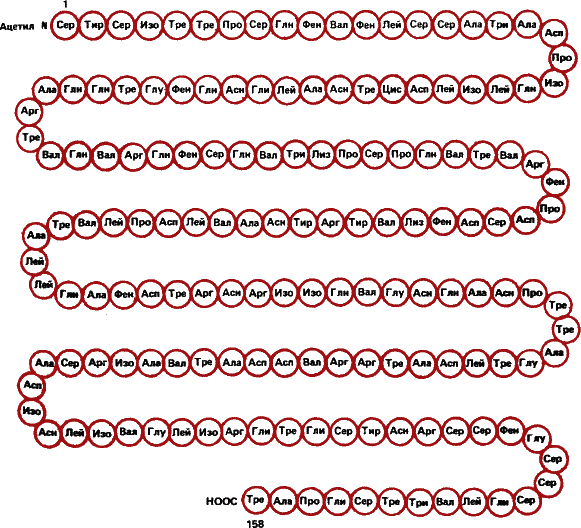
Рис. 7. Вирус табачной мозаики – первый вирус, который был описан и затем выделен в чистом виде. Вверху слева – схема строения (размеры здесь и далее даются в ангстремах – стомиллионных долях сантиметра). Справа – фотография модели вируса – однонитчатая спираль РНК в шубе из одинаковых молекул белка. Каждую такую белковую молекулу слагают в определенной последовательности 158 аминокислот.
Рассмотрим справедливость нашей первой аксиомы на ряде конкретных примеров. Итак, фенотип не может воспроизводиться без генотипа, и наоборот. Фенотип возникает по программе, кодированной в генотипе, и заодно копирует генотип для будущего поколения. Но возможны случаи, когда генотип копируется при посредстве чужого фенотипа. Тогда порог сложности, определенный фон Нейманом, снимается: возникают простейшие образования – вирусы, на примере которых можно проследить все стадии редукции фенотипа.

Рис. 8 Электронная микрофотография одного из сложных бактериальных вирусов – фага Т2. Хорошо видны молекулы белка, слагающие хвост и голову, а также хвостовые нити (слева). Размеры здесь также в ангстремах.
Самые сложные вирусы – бактериофаги, или просто фаги – паразиты бактерий. Примером могут служить так называемые Т‑четные фаги бактерии кишечной палочки. Фаг Т4напоминает по форме ручную гранату. Головка фаговой частицы – вытянутый двадцатигранник, сложенный из молекул белка, в ней содержится в компактно уложенном состоянии длинная молекула ДНК. Фаг Т4– имеет сложный механизм для вспрыскивания своего генотипа внутрь бактериальной клетки – хвостовую часть. Хвостовая часть состоит (от головки к концу) из воротничка, собственно хвоста, одетого сократимым, как мускульное волокно, чехлом, и базальной пластинки, от которой отходят шесть нитей. Нитями и зубцами базальной пластинки фаг прикрепляется к клеточной оболочке бактерии. Молекула специфического белка лизоцима на базальной пластинке разъедает оболочку бактерии, хвостовой чехол сокращается, и нить ДНК с огромной скоростью выталкивается, буквально выстреливается в цитоплазму бактерии.
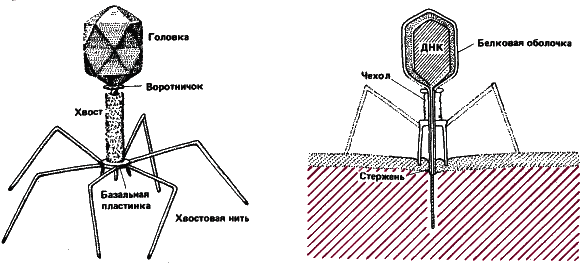
Рис. 9. Подробная схема Т‑четного фага; справа показано, как он заражает бактерию. Сначала фаг прикрепляется базальной пластинкой, опираясь при этом на хвостовые нити. Молекула лизоцима на базальной пластинке разъедает стенку бактериальной клетки, хвост сокращается, и нить ДНК выстреливается в бактерию. Фенотип фага после этого отбрасывается, он уже не нужен.
Пустая белковая оболочка – фенотип фага – уже не нужна, она отбрасывается. Эти фаги без нуклеиновой начинки образно называют тенями. Действительно, это всего лишь тени вирусов: они могут, как и живые, прикрепляться к поверхности бактерий и проедать своим лизоцимом в их оболочке отверстия. Они не могут лишь размножаться. «Голая» ДНК в бактерии начинает свою разрушительную деятельность. Прежде всего, за какую‑то минуту подавляется синтез бактериальных белков. Через пять минут после заражения белоксинтезирующий аппарат бактерии синтезирует по программе фага ферменты, размножающие фаговую ДНК. Еще через три минуты начинается синтез фаговых белков, формирующих головки и хвосты новых особей фага. Первый новый фаг при температуре 37° появляется через 13 минут, еще через 12 в бактерии их уже свыше 200. Насинтезированный за это время лизоцим разъедает стенки бактерии изнутри, и новые фаговые частицы выходят наружу, готовые заражать другие бактериальные клетки.
Рис 10. Любопытный феномен – дефектные фаги, фаги якобы без генотипа. ДНК фага, войдя в бактерию, может не взорвать ее размножением, а встроиться в бактериальную хромосому и размножаться вместе с ней. Фенотип фага тогда не образуется, фаговая ДНК обходится чужим. Такой фаг, латентный, может вернуться к активному состоянию, если культуру бактерий, зараженных им, облучить ультрафиолетом. Иногда активируется не весь генотип фага, а лишь та часть, которая ответственна за образование хвоста. Тогда в бактерии штампуются сотни хвостов, проедающих ее стенки и выходящих наружу. Они могут убивать бактерий, проедая лизоцимом их стенки, но не размножаться (ведь у них нет головки и в головке ДНК – генотип остался в бактериальной клетке). Дефектные фаги – пиоцины – размножаться сами не могут. Фенотип без генотипа бессилен. Строго говоря, нельзя говорить, что пиоцин не имеет генотипа – он просто держит его в другом месте.
Генотип таких фагов довольно сложен (около ста генов). Но есть и более простые фаги, например φX174, он гораздо меньше Т4и представляет правильный двадцатигранник без хвостовой части. Его генотип кодирует всего девять белков.
Еще проще фаг MS2: его генотип состоит из однонитчатой РНК, а фенотип лишь из трех белков: белка‑оболочки, белка катализирующего синтез РНК, и так называемого А–белка. А вот у вируса, вызывающего веретеновидность клубней картофеля, нет даже белковой оболочки. Но, попадая в клетки картофеля, его РНК строит два белка, чужеродные для растения‑хозяина.


Рис. 11. Если бактериофаги самые сложные вирусы, то так называемые вироиды – наиболее простые. Лучше всего изучен вироид, вызывающий веретеновидность клубней картофеля. Пораженные им растения легко отличить от здоровых. У них листья прижаты к стеблям, а клубни имеют характерную форму с многочисленными глазками. У вироидов нет оболочки: это голые РНК, образующие кольца; кольца, как правило, как бы сплюснуты в палочку. Этот вироид состоит из некодирующей белков РНК, в которой всего 359 нуклеотидных звеньев. Полагают, что информация о синтезе белков, появляющихся в клетках зараженного картофеля, содержится в геноме самого картофеля, но обычно заблокирована. Вироид как бы включает ее и тем самым изменяет внешний облик хозяина.
Встроенные в хромосомы хозяев вирусы могут, по‑видимому, совсем отказаться от своего фенотипа и обходиться при репликации хозяйским.
Рассмотрим теперь обратные примеры. Можем ли мы назвать живыми организмы или хотя бы клетки без генетической программы? Таковы безъядерные клетки – эритроциты млекопитающих или же клетки хрусталика глаза. Эритроцит теряет основное свойство жизни, он не поддерживает свою структуру, распадаясь за четыре месяца, и не может размножаться делением. Это всего лишь часть структуры живого организма. Сам по себе он не может жить, как любая отдельно взятая часть самолета не может летать.
На безъядерных клетках – как появляющихся в результате естественных процессов, так и получаемых в эксперименте – следует несколько задержаться: более наглядных примеров для жизни генетических программ мы не найдем.
Возьмем, к примеру, глаз млекопитающего. Как он формируется во время индивидуального развития животного? Сначала появляется так называемый глазной пузырь, затем наружный слой клеток, эктодерма, над глазным пузырем утолщается и втягивается внутрь полости будущего глаза, отшнуровывается и замыкается в пузырек. Это станет потом хрусталиком. Его стенки сложены клетками эпителия. В принципе они такие же, как те, что выстилают, скажем, внутреннюю поверхность наших кровеносных сосудов или слизистые оболочки. У них нормальные ядра, они синтезируют много белков, могут размножаться делением. Но в хрусталиковом пузырьке они меняются, постепенно переходят в длинные стекловидные прозрачные хрусталиковые волокна. Набор белков, синтезируемых ими обедняется, нарабатываются только специфичные структурные белки – кристаллины. Их три: альфа‑, бета– и гамма‑кристаллины (у птиц и пресмыкающихся синтезируется еще один – четвертый). В начале их синтеза клетка еще способна делиться. Но постепенно ее цитоплазма заполняется хрящеподобной кристаллиновой массой, сама клетка вытягивается в волокно. Параллельно идет быстрая потеря ДНК в ядре: если в начале этого процесса количество ДНК такое же, как у нормальной клетки с двойным (диплоидным) набором хромосом, то затем в хрусталиковом волокне ее не удается обнаружить самыми чувствительными методами. Это волокно уже не делится и не поддерживает свою структуру. Тем самым оно отличается от многих других клеток организма, в норме хотя и не делящихся, но сохраняющих ядро – такие клетки еще способны к «ремонтным работам».
Удивительно сходная картина наблюдается при образовании эритроцитов, хотя конечные стадии процессов совершенно различны. Так называемые стволовые клетки кроветворной системы могут делиться практически неограниченное число раз. Но часть из них превращается в проэритобласты: в них уже начинается синтез той РНК, которая послужит потом матрицей для синтеза гемоглобина. На этой стадии ядра еще нормальны и клетки способны к делению. При переходе в следующую стадию (на этой стадии эритобласты могут окрашиваться разными красками и поэтому получили название полихроматофильных) ядра теряют активность.
У рыб, земноводных, пресмыкающихся и птиц ненужное, в общем‑то, ядро так и остается в зрелом эритроците.
У млекопитающих, а стало быть, и у людей процесс идет дальше – ядро исчезает. Такая клетка называется ретикулоцитом. У лабораторных животных, например, кроликов, специфическими ядами можно остановить процесс кроветворения на этой стадии. Тогда в костном мозге накапливаются ретикулоциты. Для биохимика эти клетки – сущий клад! В них нет ядер, но много матричных РНК, белоксинтезирующий аппарат работает и гонит лишь одни гемоглобины. Но вот будущий эритроцит насыщен гемоглобином до предела. Белоксинтезирующий аппарат редуцируется, снижает активность, а зрелый эритроцит, войдя в сосуды, начинает свою жизнь – в среднем 4‑месячную.
Клетки без генетической программы, без ядра (энуклеированные) можно получить искусственно. Эмбриологи давно уже научились проводить такие операции на оплодотворенных яйцеклетках некоторых животных: их сначала откручивают на центрифуге, отчего ядро как более тяжелое смещается в нижнюю часть клетки, а уж после этого при определенном навыке относительно нетрудно разделить под микроскопом яйцеклетку на часть без ядра и на часть с ядром. Результат всегда один: безъядерная часть постепенно рассасывается, из части с ядром развивается нормальный организм.
Но наиболее удачный объект для подобных опытов – обитающая в теплых морях, включая Черное море, водоросль ацетабулярия. Строение ее очень характерно. Так, например, распространенная у нас ацетабулярия средиземноморская имеет вид миниатюрной – несколько миллиметров и более – поганки с плоской шляпкой на длинной ножке, шляпка у некоторых видов достигает 5 сантиметров в диаметре. И этот грибок состоит из одной клетки! Оперировать ее можно без особых ухищрений, тем более что ядро этой водоросли помещается в самой нижней части ножки, у корнеобразных выростов ризоидов, которыми водоросль прикрепляется к грунту. Хирургические операции сводятся к тому, что водоросль разрезают на куски и наблюдают за их дальнейшим «поведением».
Оказалось, что если рассечь ножку, то шляпка, в конце концов, погибнет, хотя некоторое время она еще может, используя энергию солнечного света, синтезировать органику из углекислого газа и воды. А нижняя часть ножки, где находится ядро, снова обзаведется шляпкой – все, как у ящериц. Но самое любопытное дальше: когда для восстановившей себя ацетабулярии приходит время делиться, то шляпка достается лишь одной половинке, одной новой особи, другая обязана «строить» себе другую шляпку.
Но и это еще не все. Разные виды ацетабулярий отличаются друг от друга главным образом строением шляпки, зонтика. У средиземноморской, например, зонтик круглый и вогнутый, а у ацетабулярии Веттштейна он рассечен на лопасти и по форме похож на цветок. Был проведен такой эксперимент: ножку без шляпки от средиземноморской ацетабулярии пересадили на ризоиды ацетабулярии Веттштейна. Получился «вегетативный гибрид». Он быстро надстроил себе шляпку, и она оказалась рассеченной на лопасти. Впрочем, удивительного тут ничего нет: ведь ядро, а значит, и генетическая программа в ней, в этой химере, были от веттштейновской водоросли.
Все эти примеры о великой роли генетических программ относятся к клеточной форме организации живого. А как обстоит дело у неклеточных форм жизни, у вирусов?
Открытый первым и наиболее хорошо изученный вирус табачной мозаики (ВТМ) – длинная палочка, точнее трубка, состоящая на 95 процентов из белка и 5 процентов РНК. Трубчатый белковый чехол состоит примерно из 2300 молекул белка; на внутренней его поверхности пологой спиралью расположена длинная нить РНК.
В уксусной кислоте (66 процентов) ВТМ распадается на отдельные молекулы белка и РНК. Если кислоту нейтрализовать щелочью, молекулы белка снова слипаются, образуя длинные трубчатые гильзы. РНК тут не обязательна, в ее отсутствие образуются столь же длинные, а то и длиннее обычных, белковые трубки, внешне неотличимые от исходных вирусных частиц.
Но заразить клетку табака они не могут. Основное свойство живого – самовоспроизведение – утеряно вместе с генетической программой.
Наш анализ первой аксиомы завершим примером, который хочется назвать «История со скрепи». Вирусологов последние двадцать лет весьма интересовала загадочная болезнь овец – инфекционная чесотка, поражающая периферийные нервные окончания, развивающаяся очень медленно и в конце концов приводящая к смерти. Ее назвали скрепи (scrape); она очень напоминала другие болезни овец (висна, рида, мэди) и людей (амиотрофический боковой склероз). Известно уже около 15 таких болезней, так называемых медленных вирусных инфекций. Из человеческих, пожалуй, наиболее экзотична куру – «хохочущая смерть», до недавнего времени поражавшая папуасов новогнинейского племени форе. Здесь экзотичен способ заражения: согласно религиозным обычаям форе на поминках по родствевнику еще несколько лет назад обязаны были съедать его мозг. А куру, как и подавляющее большинство медленных вирусных инфекций, тяготеет к клеткам нервной системы.
Парадоксальность положения заключается в том, что электронный микроскоп тут бессилен. Возбудителя скрепи или куру в него никак не удавалось разглядеть. Вирусологи оказались в положении Луи Пастера – тот мог судить, содержится ли вирус бешенства в мозговой ткани, только заразив последней подопытное животное. И самое главное, по некоторым данным частицы скрепи не содержали нуклеиновых кислот: ни ДНК, ни РНК! Это почему‑то приводило в восторг антидарвинистов. Согласно их логике, если материальным субстратом наследственности может быть другое вещество, кроме нуклеиновых кислот, то это опровергает теорию эволюции Дарвина. На мой взгляд, это утверждение равносильно следующему: если стихи можно написать на чем‑либо, кроме бумаги, то поэзии не существует.
Оставим антидарвинистов и задумаемся: каким же образом вирус скрепи размножается, воспроизводит самого себя без генотипа?
Несколько лет назад автор этой книги был в экспедиции на юге Туркмении. Мы сидели на заброшенном в песках кордоне, днем ловили змей и ящериц, а вечером, когда жара спадала, за зеленым чаем вели длинные разговоры обо всем, включая молекулярную биологию. Не миновали мы и скрепи. Как истолковать отсутствие в этом вирусе нуклеиновых кислот? Да и вирус ли это?
Тогда на досуге я придумал довольно сложную схему, позволявшую скрепи размножаться на базе генотипа хозяина. В общем, получалось что‑то вроде короткого замыкания генетического аппарата клетки. Эта спекуляция одним ударом объясняла и медленное развитие инфекции, и приуроченность вируса к неделящимся клеткам нервной системы. Моему коллеге она понравилась, и он даже посоветовал ее опубликовать. Я не послушался совета и правильно сделал. Финал истории прост: недавно в вирусе скрепи обнаружили нуклеиновую кислоту.
Первую аксиому биологии не понадобилось защищать – она сама постояла за себя.
Итак, мы сформулировали первое положение, лежащее в основе всей живой материи, только для нее характерное и общее для всех живых субъектов – от примитивнейшего вируса до человека.
Однако из него естественно вытекает несколько проблем, решение которых приводит к формулировке биологической аксиомы № 2. Среди них главные:
1. Достаточно ли информации, содержащейся в генотипе, для набора инструкций, потребного для построения фенотипа? Вопрос можно сформулировать и иначе: что сложнее – генотип или фенотип? Сколько информации потребуется для описания того и другого?
2. Каким образом из поколения в поколение воспроизводится генетическая программа, по которой развиваются организмы?
Переходим ко второй аксиоме биологии.
И снова немного истории. В 1927 роду на III Всесоюзном съезде зоологов, анатомов и гистологов в Ленинграде наш блестящий биолог Николай Константинович Кольцов сделал доклад, в котором впервые была четко сформулирована вторая аксиома биологии. Принцип Кольцова до сих пор остается незыблемым, несмотря на то, что наши представления о природе наследственных молекул совершенно изменились.
В начале доклада Кольцов припомнил давнее событие – Московский съезд естествоиспытателей и врачей, состоявшийся в 1893 году. Два тогдашних выступления особенно врезались в его память, тогда молодого исследователя, изучавшего анатомию лягушки.
Профессор М. А. Мензбир рассказал о нашумевших идеях Августа Вейсмана, разделившего организм на наследственную плазму и сому (аналоги сегодняшних генотипа и фенотипа). Из теории Вейсмана следовало, что генотип располагается в клеточном ядре и передается от поколения в поколение яйцеклетками и спермиями.
И на том же съезде химик А. А. Колли путем простейших математических выкладок, основываясь на далеко еще и во многом неверных тогдашних представлениях о природе белков, показал, что в головке спермия может уместиться очень мало белковых молекул: несколько десятков, то есть примерно столько же, что и хромосом.
Странным образом никто тогда, кроме Кольцова, не сопоставил оба этих выступления. Да и сам Николай Константинович вынес свои идеи на всеобщее обсуждение только после более чем тридцатилетних размышлений, уже после того, как родилась на свет генетика Моргана и белковая химия шагнула далеко вперед.
Вывод его был прост хромосома – это гигантская молекула. Впоследствии, в 1935 году он назвал хромосомы «наследственными молекулами».
Согласно Кольцову в хромосоме в линейной последовательности располагаются белковые молекулы – гены. Каждый ген – цепочка аминокислотных остатков, соединенных пептидными связями.
Рис. 12. Таким представлял процесс деления хромосомы Н. К. Кольцов, гениально предугадавший необходимость для жизни матричного синтеза.
Напомним, кстати, читателям, что такое аминокислота. Этого названия заслуживает любое соединение, содержащее одновременно аминогруппу – NH2и радикал органических кислот – COOH. Пептидная связь возникает между этими группировками: при этом отщепляется молекула воды. Белки состоят из сотен и тысяч аминокислотных остатков, соединенных пептидными связями. Кольцов предположил, что все наследуемые свойства организмов закодированы в хромосомах порядком чередования разнообразных аминокислотных остатков.
Но отсюда следовало, что заново возникать подобные молекулы не могут. Слишком мала вероятность того, что аминокислоты сами по себе, без какого‑нибудь упорядочивающего фактора соберутся в нужную последовательность. А ведь она воспроизводится в каждом поколении и вероятность ошибки ничтожна. Кольцов приводил пример с цепочкой всего из 17 аминокислот, возможно существование триллиона вариантов таких цепочек, различающихся чередованием остатков! Но такая цепочка (гептакайдекапептид) гораздо проще большинства природных белков.
Теперь, когда последовательность аминокислот известна для многих десятков, если не сотен, белков, можно привести еще более убедительный пример, как это делает Манфред Эйген[4]. Цитохром Сне самый большой белок, в нем лишь около сотни аминокислотных остатков. Эйген подсчитал, что число вариантов такой последовательности около 10130(единица со ста тридцатью нулями). Трудно представить столь огромную величину. Если бы вся Вселенная (все планеты, звезды и галактики) состояла из цитохрома С, в ней могло уместиться только около 1074молекул! Это ли не подтверждение мысли Кольцова!
И Кольцов делает следующий вывод: