Организация эу- и гетерохроматина. Структура и химия хроматина.
В ядре сосредоточена большая часть ДНК эукариотической клетки - 90%.в составе хромосом.Материал хромосом - совокупность глыбок, зерен и волоконец – хроматина.
Химический состав хроматина (хромосом) эукариотической клетки
Большая часть объема хромосом представлена ДНК и белками. Заметные химические компоненты хромосом - РНК и липиды. Среди белков (65% массы хромосом) выделяют гистоновые (60-80%) и не-гистоновые. Также присутствуют полисахариды, ионы металлов (Ca, Mg) и др. Особое место среди хромосомных белков принадлежит гистонам. В составе нуклеогистонового комплексаДНК менее доступна ферментам нуклеазам, вызывающим ее гидролиз (функция защиты). Гистоны выполняют структурную функцию, участвуя в процессе компактизации хроматина. Гистоновые белки представлены пятью видами (фракциями): Н1, Н2А, Н2В, Н3 и Н4.
Число ядерных негистоновых белковпревышает несколько сотен. Они удерживают «открытую» конфигурацию хроматина, «разрешающую» доступ к биоинформации ДНК, то есть ее транскрипцию.
К категории «временных» относятся цитозольные белки-рецепторы (функционально-транскрипционные факторы), захватывающие сигнальные молекулы, в комплексе с которыми они проникают в ядро и их активируют.
РНК хромосомпредставлена продуктами транскрипции, еще не покинувшими место синтеза, - непосредственный продукт транскрипции генов или пре-и(м)РНК, пре-рРНК, пре-тРНК транскрипты. Некоторые виды РНК «временного внутриядерного пребывания» создают условия для основного процесса, выполняя сигнальную функцию. Так, репликация ДНК требует для своего начала образуемой «на месте» РНК-затравки (РНК-праймер), которая по завершении процесса разрушается здесь же в ядре.
В зависимости от степени компактизации материал интерфазных хромосом представлен эухроматином и гетерохроматином . Эухроматин - низкая степень компактизации и неплотная «упаковка» хромосомного материала. Эухроматин представлен, в основном, ДНК с уникальными последовательностями нуклеотидов. Гены из эухроматизированного участка хромосомы, оказавшись в гетерохроматизированномучастке или рядом с ним, обычно инактивируются.
Гетерохроматинотличается высокой степенью компактизации, то есть плотной «упаковкой» материала хромосомы. Большая его часть представлена умеренно или многократно повторяющимися нуклеотидными последовательностями ДНК. К первым относятся мультикопийные гены гистонов, рибосомных и транспортных РНК.
58. Уровни структурной организации хроматина. Компактизация хроматина.
На протяжении клеточного цикла хромосома сохраняет структурную целостность благодаря компактизации-декомпактизации (конденсация-деконденсация)хромосомного материала – хроматина. Вследствие компактизации при переходе хромосом из интерфазной формы в митотическую суммарный линейный показатель сокращается примерно в 7-10 тыс. раз.
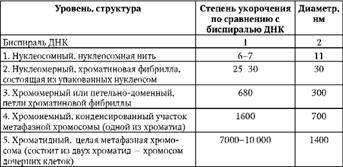
Таблица 2.1.Последовательные уровни компактизации хроматина.
В образовании нуклеосомной нити ведущая роль принадлежит гистонам Н2А, Н2В, Н3 и Н4. Они образуют белковые тела или коры,состоящие из восьми молекул. Молекула ДНК комплексуется с белковыми корами, спирально накручиваясь на них-биспирали. Свободную от контакта с корами ДНК наз линкерной(связующая).Отрезок ДНК + белк кор = нуклеосома. Благодаря нуклеосомам в промоторных участках ДНК заблокированы области инициации (начала) транскрипции. Для того чтобы инициаторный комплекс возник, нуклеосомы должны быть «вытеснены» из соответствующих фрагментов ДНК.
Образование хроматиновой фибриллы диаметром 30 нм (второй уровень компактизации) происходит с участием гистона Н1, который, связываясь с линкерной ДНК, скручивает нуклеосомную нить в спираль.
На следующем петельно-доменном -укладка фибриллы диаметром 30 нм в петли. В этом процессе активная роль отводится негистоновым белкам. Основания петель «заякорены» в ядерном матриксе. Петля содержит от одного до нескольких генов(петельный домен).
На следующем уровне компактизации «сложенные» фибриллы превращаются в метафазные хроматиды (хромосомы будущих дочерних клеток).
Максимальная степень компактизации достигается на пятом уровне в структурах, известных как метафазные хромосомы с диаметром 1400 нм. Такая структура обеспечивает оптимальное решение задачи транспортировки генетического материала в дочерние клетки в анафазе митоза.