Лекция №2
Темная окраска выполняет защитную функцию на темном фоне, светлая — на светлом. К середине XX в. меланизм был выявлен как у 70 видов бабочек в различных странах, так и у других насекомых, например, в ленинградских популяциях упоминавшейся двухточечной коровки (JI. 3. Кайданов, 1996).
Р. Келли с сотрудниками (R. Kelly et al., 2002) изучали генетическую структуру ручейника Wormaldia taganunana, эндемика Канарских островов, определяя аллозимную изменчивость 11 локусов в 5 из 8 популяций. Сравнивался поток генов, генетическая изменчивость и генетическая популяционная структура этого вида с таковыми более широко распространенных видов ручейников, в частности канарских популяций неэндемика Канарских островов Mesophylax asperses. Было показано, что ограниченный ареал распространения W. tagananana отчасти связан с относительно меньшей генетической изменчивостью и более низким потоком генов в популяциях W. tagananana.
Рыбы. Корейские популяции медаки Oryzias latipes состоят из двух четких групп, в которых по изменчивости выявились 13 аллоферментных локусов: восточно-корейские и китайские/западнокорейские. На западном побережье обе группы распространяются мозаично (T.Yusuke, 2004).
Для внутривидовых сопоставлений природных популяций нельмы и искусственно поддерживаемой популяции белорыбицы учитывались частоты генов по результатам биохимической обработки 23 — 25 изоферментных локусов (Т. С. Голованова, 2004). Для исследованных выборок белорыбицы и нельмы были рассчитаны основные параметры генетической изменчивости — величины уровня полиморфизма и средней ожидаемой гетерозиготности (табл. 7). Уровень полиморфизма, средняя гетерозиготность и среднее число аллелей на локус у нельмы и белорыбицы имели сходные показатели.
Показатели генетического разнообразия исследованных выборок белорыбицы и нельмы приведены в табл. 8. Показатели доли межпопуляционного разнообразия FST для нельмы и белорыбицы различны. Доля генетического разнообразия, приходящегося на различия между выборками нельмы, выше, чем доля, приходящаяся на различия между выборками белорыбицы, взятыми в разных местах и в разное время хода на нерест.
Таблица 7. Уровень полиморфизма, среднее число аллелей на локус и средняя ожидаемая гетерозиготность в популяциях нельмы и белорыбицы (по 23 локусам) (по Т. С. Головановой, 2004)
'Локус считается полиморфным, если частота основной аллели не превышает 0,99. 2Локус считается полиморфным, если частота основной аллели не превышает 0,95. |
Таблица 8. Показатели генетического разнообразия для белорыбицы и нельмы (по полиморфным локусам) (по Т. С. Головановой, 2004)
|
| 1FSt — доля межпопуляционного разнообразия. 2Nem — число мигрантов на поколение. |
Доля генетического разнообразия, приходящаяся на различия между выборками нельмы р. Северной Двины и оз. Кубенское (FST = 0,0032), гораздо ниже, чем доля, приходящаяся на различия между выборками нельмы р. Оби и р. Печоры (FST = 0,1294). Средний уровень генетической миграции Nem, оцененный на основе стандартизированных генетических варианс по аллозимным локусам, в среднем соответствует таковым, отмеченным для лососевых и сиговых видов рыб.
На основании полученных данных сделан вывод об однородности проанализированных выборок: ни по одному из исследованных локусов не было обнаружено статистически достоверных различий. Эти результаты подтверждают точку зрения М.А.Летичевского (1960), что волжская белорыбица образует единое стадо с растянутым на несколько месяцев временем хода на нерест (Т. С. Голованова, 2004). Снижение генетического разнообразия у молоди белорыбицы выпуска 2001 и 2002 гг. не повторилось в 2003 г., когда для искусственного воспроизводства использовалось более 100 производителей.
Амфибии и рептилии. В. А. Межжерин (1997) проанализировал генетическую изменчивость озерной лягушки из Восточного Казахстана по 32 биохимическим локусам 13 ферментов. Из них 4 локуса оказались полиморфными, тогда как в европейских популяциях они были исключительно мономорфны. Гетерозиготность популяций из Учарала (Талды-Курганская область) составляла 3,6%, из Алма-Аты — 10,9%.
Хороший пример последствий изучения генетической изменчивости дает обыкновенная чесночница Pelobates fuscus. Согласно размеру генома, имеются две отличные группы в пределах ареала этого вида в Восточной Европе (L. Y. a. Borkin et al., 2003). Географически животные с меньшим размером генома распределены на западе (средний размер генома 8,83 пкг), а с большим (9,35 пкг) — на востоке (рис. 20).
 Рис. 20. Размер генома выборок обыкновенной чесночницы (по L. Ya. Borkin et al., 2003, с изменениями)
Рис. 20. Размер генома выборок обыкновенной чесночницы (по L. Ya. Borkin et al., 2003, с изменениями)
|
Западный тип обыкновенной чесночницы был зарегистрирован в Латвии, Белоруссии, Украине (Сумская, Черниговская, Киевская и Одесская области), Молдавии и на западе европейской части России (Калининградская, Псковская, Ленинградская, Ярославская, Московская и Тульская области). Восточный тип зафиксирован в Харьковской области Украины, южных, центральных и восточных областях европейской части России, а также в северо-западной части Казахстана. Таким образом, восточный тип обыкновенной чесночницы характерен для бассейна Средней и Нижней Волги, Центрально-Черноземного района России и Ставропольского края.
Кроме генома некоторые выборки были также исследованы с применением метода аллозимного анализа. Данные по белкам ясно указали на присутствие двух главных групп среди популяций обыкновенной чесночницы из Восточной Европы. Географически, как правило, одна группа состояла из западных выборок, в то время как другая группа включала восточные выборки. Картирование ал- лозимных данных показало явную географическую неоднородность выборок. Вообще с запада на восток аллели, характерные для западного типа, имели тенденцию к замене таковых восточным типом. Однако оказалось, что частота «восточных» аллелей была весьма высока в некоторых западных выборках, идентифицированных проточной ДНК-цитометрией, и, наоборот, «западные» аллели были зарегистрированы в выборках восточного типа (рис. 21).
На основе этих данных авторы исследований (Л.Я.Боркин и др., 2004) пришли к выводу о том, что обыкновенная чесночница представлена в Восточной Европе двумя криптическими (т. е. морфологически сходными) формами, таксономический статус которых пока не ясен, но, возможно, что их ранг не меньше видового.
Молекулярно-генетическая дифференциация средиземноморской черепахи, полученная при использовании 213 ПЦР-маркеров, соответствует выделению ранее описанных подвидовых групп (С. К. Семенова и др., 2004). Черепахи, собранные из разных районов Дагестана, представляют собой более полиморфную и гетерогенную группу по сравнению с черепахами из разных районов Средней Азии.
Млекопитающие. Р. Силандер с сотрудниками обследовали 30 популяций белоногого хомячка Peromyscus polionotus на побережье Мексиканского залива. Средняя гетерозиготность популяций п-ова Флорида на 75 % превосходила этот показатель, отражающий основную часть популяций ареала. Низкой гетерозиготностью отличались островные популяции западного побережья, населя-
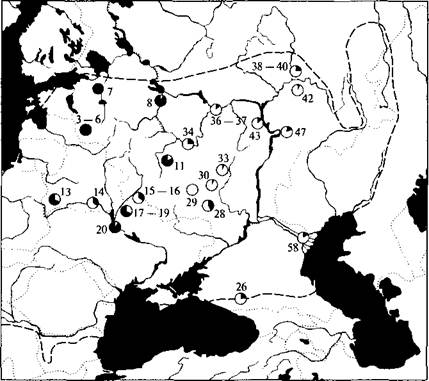 Рис. 21. Распределение «западных» (черный цвет в кружках) и «восточных» (белый цвет в кружках) аллелей в двух криптических формах обыкновенной чесночницы из Восточной Европы (по L. Ya. Borkin et al., 2003). Пунктиром обозначена граница ареала обыкновенной чесночницы, точечными линиями — границы государств
Рис. 21. Распределение «западных» (черный цвет в кружках) и «восточных» (белый цвет в кружках) аллелей в двух криптических формах обыкновенной чесночницы из Восточной Европы (по L. Ya. Borkin et al., 2003). Пунктиром обозначена граница ареала обыкновенной чесночницы, точечными линиями — границы государств
|
Л.А. Балмышева (2001) исследовала полиморфизм митохондриальной ДНК соболя из четырех популяций, обитающих на Камчатке, Сахалине, в Хабаровском крае и Магаданской области. В результате рестрикционного анализа гена цитохрома b выявлены три дифференцированные формы — гаплотипы А, В и С, которые распределялись своеобразным образом и с определенной частотой. Это позволило сделать вывод, что на северо-востоке России обитает два подвида соболя. В популяциях лесных полевок близ Магадана была обнаружена определенная смена генотипов в ходе естественной флуктуации группировок (С. В. Курышев, Ф. Б. Чернявский, 1988).
Более подробные сведения о генетической изменчивости популяций различных видов растений и животных, о генетике популяций и т.п. можно почерпнуть в трудах Ф.Добжанского, Н. П. Дубинина, А. В.Яблокова и многих других авторов, в монографиях и учебниках JT. Меттлера и Т. Грегга (1972), Ф.Айалы (1984), Ю. П.Алтухова (1989), М. М.Тихомировой (1990), Л.З.Кай- данова (1996), Ф. Хедрика (2003) и др.
Контрольные вопросы
1. Какие показатели популяций называются статическими, а какие — динамическими? В чем их разница? 2. Чем характеризуется половая структура популяций? 3. Каковы особенности первичного, вторичного и третичного соотношения полов у отдельных растений и животных? 4. Что такое арренотокия и цикломорфоз? 5. Какие условия могут повлиять на соотношение полов в популяции? 6. В чем сущность протандрии и прото- гинии? 7. Дайте определение поколению, приплоду и возрастной группе.
8. В чем сущность построения возрастных пирамид? Приведите примеры.
9. Какие возможны соотношения отдельных возрастных категорий?
Каковы варианты пространственного размещения особей? Приведите примеры. 11. Что такое радиус репродуктивной активности? 12. В чем отличие оседлых животных от кочевых? 13. В чем преимущества группового образа жизни? В чем выражается эффект группы? 14. Охарактеризуйте типы интеграции индивидуумов. 15. Дайте характеристику колонии, семье, стае. 16. В чем разница стад с вожаком и лидером? 17. Что такое генетическая гетерогенность и генетический полиморфизм? 18. В чем особенности генетической структуры отдельных растений и животных?