Виды РНК и их строение
В клетках организмов содержатся три вида РНК: рибосомная (рРНК), информационная или матричная (mРНК) и транспортная (тРНК). Основная функция РНК – перенос генетической информации от ДНК на белки. Первые сведения об участии РНК в синтезе белков были получены в 1941-42 г.г. Однако более полное представление о строении и функциях каждого из трёх видов РНК было достигнуто в 1956-1961 г.г., когда были открыты рибосомы и рибосомная РНК, а также выяснена биологическая роль информационной и транспортной РНК.
Рибосомная РНК локализована в составе рибосом цитоплазмы и внутриклеточных органелл – митохондрий и хлоропластов. Однако её синтез происходит в ядре. У высших организмов местом синтеза большей части рРНК является ядрышко. Гены рРНК в хромосомах клеток представлены многими повторяющимися копиями. Полинуклеотидные последовательности рРНК содержат информацию о структуре рибосомных белков, многие из которых после их синтеза образуют с рРНК комплексы и химические соединения в виде нуклеопротеидов. В количественном отношении рРНК преобладает в клетке над другими видами РНК, на её долю приходится до 90% от общего содержания РНК в клетке.
В клетках прокариот каждая рибосома содержит три молекулы рРНК, различающиеся по составу нуклеотидов и молекулярным массам. Большая по размерам субъединица рибосомы прокариот ( 50S-субъ-единица) содержит 23S-РНК, состоящую из 3300 нуклеотидных остатков, и 5S-РНК, включающую 120 нуклеотидных единиц. В малой субъединице находится 16S-РНК, в полинуклеотидной цепи которой насчитывается 1700 нуклеотидных остатков.
В цитоплазматических рибосомах высших организмов содержатся четыре вида РНК: 28S-РНК, имеющие в своём составе 5000 нуклеотидных остатков (н.о.); 18S-РНК (2000 н.о.), 5,8S-РНК (150 н.о.) и 5S-РНК (120 н.о.). Гены, кодирующие структуру 28S-РНК, 18S-РНК и 5,8S-РНК локализованы в ядрышке, а генетическая информация о структуре 5S-РНК содержится в других участках хромосом вне ядрышка. В большой субъеденице цитоплазмотических рибосом высших организмов (60S-субъединица) локализованы 28S-РНК, 5,8S-РНК и 5S-РНК, в малой субъединице (40S-субъединица) – 18S-РНК. В хлоропластных рибосомах содержатся такие же по размерам молекулы рРНК, как и клетках прокариот. Митохондриальные рРНК по размерам и структуре также находятся ближе к бактериальным.
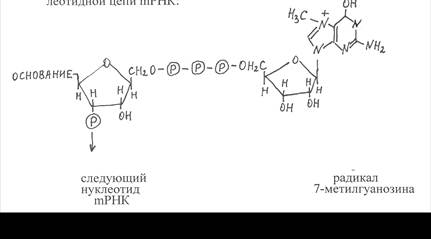 |
Матричная РНК. Матричная, или информационная, РНК в отличие от рРНК является короткоживущей формой РНК. Период полужизни её молекул у бактерий составляет в среднем 2-3 минуты, а у высших организмов – от нескольких часов до 1-2 суток. Основная функция mРНК – копирование нуклеотидных последовательностей ДНК, кодирующих структуру определённых белковых полипептидов, и участие совместно с рибосомами и тРНК в синтезе белков. В клетках прокариот синтезируемая mРНК содержит нуклеотидные последовательности одного или нескольких генов. В клетках эукариот каждая молекула mРНК копирует структуру одного гена и обычно включает 1500-2000 нуклеотидов. На её 5'-конце находится радикал 7-метилгуанозина, а к 3'-концу присоединяется полинуклеотидная цепь, включающая около 100 повторяющихся остатков адениловой кислоты (поли-А), за исключением mРНК гистоновых белков. Радикал 7-метилгуанозина соединяется с 5'-концом mРНК трифосфатной группировкой, которая образует эфирные связи с остатками рибозы 7-метилгуанозина и последующего остатка нуклеотида на 5'-конце полинуклеотидной цепи mРНК.
В клетках организмов количество mРНК обычно составляет 1-3 % от общей суммы РНК. По наличию в клетке тех или иных видов mРНК можно судить об активности функционирования соответствующих генов, кодирующих структуру определённых белков, которые в данный момент обеспечивают жизнедеятельность клетки. Молекулы рибосомной и матричной РНК образуются из одной полинуклеотидной цепи, которая на отдельных участках формирует короткие двойные спирали. Между участками двойной спирали в виде разной формы петель находятся участки одноцепочечной РНК (рис. 43). Меньшие по размеру одноцепочечные фрагменты в виде петель также довольно часто прерывают структуру двойных спиралей РНК.
Образование двойных спиралей в молекулах РНК инициируют водородные связи, возникающие между парами оснований А–У и Г–Ц, а в отдельных случаях водородными связями скрепляются также пары оснований Г–У. Цепи РНК, образующие двойные спирали, антипараллельны и существуют в правозакрученной А-форме, подобной А-форме молекул ДНК. За счёт взаимодействия элементов вторичной структуры молекулы РНК формируют довольно компактную и упорядоченную третичную структуру. Она стабилизируется образованием водородных связей между основаниями, входящими в состав одноцепочечных участков в разных частях молекулы РНК, образуя с ними комплексы в виде двойной спирали.
Одноцепочечные участки могут взаимодействовать также с основаниями в составе двойных спиралей РНК. Кроме того, возможно образование водородных связей при взаимодействии НО-групп рибозы с азотом оснований. Для стабилизации третичной структуры РНК важную роль играют также катионы Mg2+ и другие двухвалентные катионы, которые связываются с фосфатными остатками и отрицательно заряженными группировками оснований полинуклеотидной цепи.
Транспортная РНК в клетках организмов находится в растворимом состоянии в жидкой фазе и осуществляет функцию избирательного связывания амминокислот и переноса их к рибосомам, осуществляющим синтез белков. Для каждой аминокислоты имеются свои виды тРНК. Молекулы тРНК прокариот и высших организмов по размерам и структурным характеристикам существенно не различаются. Они образуют полинуклеотидную цепь, состоящую из 70-90 нуклеотидных остатков, которая имеет пространственную структуру в виде «клеверного листа» (рис. 44).
В молекуле любой тРНК имеется четыре участка, образующих двойные спирали за счёт водородных связей между комплементарными основаниями. Три таких участка заканчиваются одноцепочечными петлями, включающими модифицированные нуклеотиды, а четвёртый участок содержит 3'- и 5'-концы полинуклеотидной цепи тРНК. На 3'-конце любой тРНК находится последовательность из трёх нуклеотидов Ц-Ц-А, служащая для связывания аминокислоты. На противоположном конце третичной структуры тРНК имеется одноцепочечная петля, содержащая в центральной её части последовательность из трёх нуклеотидных остатков, называемую антикодоном. Она комплементарна одному из участков mРНК и в ходе синтеза полипептидной цепи соединяется водородными связями с этим участком mРНК, обеспечивая таким образом включение в полипептид соответствующего аминокислотного остатка, связанного с данной тРНК. У каждой тРНК имеется свой антикодон.
 |
Характерной особенностью тРНК является наличие в их структуре большого количества модифицированных нуклеотидов. Одной из распространённых форм модификации нуклеотидов является метилирование оснований или остатков рибозы по гидроксильной группе у второго углеродного атома. В результате присоединения водорода к азотистым основаниям происходит образование дигидропроизводных различных нуклеотидов. Остатки уридиловой кислоты в ходе модификации превращаются в остатки псевдоуридиловой кислоты, в которой рибоза соединяется не с азотом, а с пятым углеродным атомом урацила:
Остаток псевдоурациловой кислоты в полинуклеотидных цепях тРНК обозначают символом ψ. Остаток урацила псевдоуридиновой кислоты, как и в составе уридиловой кислоты, образует комплементарную пару с остатком аденина, входящего в состав адениловой кислоты. В антикодоновой петле тРНК к 5'-концу антикодона всегда присоединяется остаток уридиловой кислоты (У), а со стороны 3' конца – остаткок сильно модифицированного нуклеотида, обозначаемого символом Y. Этот нуклеотид образуется из гуаниловой кислоты в результате модификации в ней азотистого основания гуанина. У некоторых тРНК в состав антикодонов наряду с остатками типичных нуклеотидов входят также остатки инозиновой кислоты.