Механизмы образования и превращений триоз, эритрозы, пентоз и гексоз.
В фотосинтезирующих клетках моносахариды синтезируются в рассмотренных нами реакциях цикла Кальвина. Промежуточными продуктами этого цикла являются 3-фосфоглицериновый альдегид, фосфодиоксиацетон, эритрозо-4-фосфат, рибозо-5-фосфат, фруктозо-6-фосфат. Фосфодиоксиацетон и 3-фосфоглицериновый альдегид образуются также в ходе реакций анаэробной стадии дыхания. Главный механизм образования указанных выше моносахаридов в нефотосинтезирующих клетках – это ракции пентозофосфатного цикла.
В 1935-38 г.г. исследованиями ряда учёных было показано, что при ингибировании основного дыхательного пути углеводов процесс их окисления не прекращается, так как инициируются реакции превращения гексоз в пентозы на уровне фосфорнокислых эфиров, которые сопровождаются окислением и отщеплением от гексоз атомов углерода в виде СО2. Такой путь окисления гексоз имеет циклический механизм и получил название пентозофосфатного пути или пентозофосфатного цикла. Как и реакции цикла Кребса, пентозофосфатный путь окисления глюкозы инициируется в присутствии кислорода.
Все ферменты, катализирующие реакции пентозофосфатного цикла, локализованы в жидкой фазе цитоплазмы, а также в хлоропластах и пропластидах. Реакции этого цикла инициируются при усилении биосинтетических процессов, связанных с потреблением восстановленных динуклеотидов НАДФ×Н. В нефотосинтезирующих клетках реакции пентозофосфатного цикла являются основным источником этих восстановленных продуктов.
Повышение концентрации окисленной формы динуклеотидов НАДФ+ вызывает активацию дегидрогеназ, катализирующих ключевые реакции пентозофосфатного цикла. Кроме того, промежуточный продукт пентозофосфатного цикла 6-фосфоглюконовая кислота ингибирует фермент фосфофруктокиназу, инициирующий реакции гликолиза, что способствует переводу дыхательного процесса на пентозофосфатный путь окисления гексоз.
|
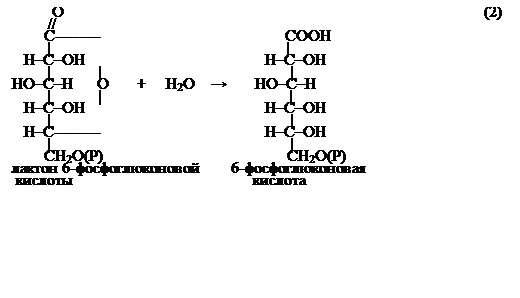
С участием фермента глюконолактоназы (3.1.1.17) лактон далее превращается в 6-фосфоглюконовую кислоту:
 |
|
Для осуществления дальнейших реакций пентозофосфатного цикла необходимо окисление в рибулозо-5-фосфат ещё одной молекулы глюкозо-6-фосфата, в связи с чем реакции 1, 2 и 3 повторяются, образуя восстановленные динуклеотиды НАДФ×Н и рибулозо-5-фосфат. После этого происходит изомеризация рибулозо-5-фосфата в ксилулозо-5-фосфат под действием фермента рибулозофосфатэпимеразы и в рибозо-5-фосфат – с участием фермента рибозофосфатизомеразы:
|
Далее вступает в действие фермент транскетолаза, который переносит концевой двууглеродный радикал с кетонной группой от ксилулозо-5-фосфата на рибозо-5-фосфат, в результате чего образуются два новых фосфата моносахаридов – седогептулозо-7-фосфат и 3-фосфоглицериновый альдегид:
|
Затем под действием фермента трансальдолазы осуществляется перенос трёхуглеродного радикала от седогептулозо-7-фосфата на 3-фосфо-глицериновый альдегид с образованием фруктозо-6-фосфата и эритрозо-4-фосфата:
|
СН₂ОН фруктозо-6-фосфат
седогептулозо-7-фосфат
Образовавшийся фруктозо-6-фосфат в следующей реакции изомеризуется в глюкозо-6-фосфат с участием фермента глюкозофосфатизомеразы:
|
|
СН2ОН Н СН2ОН Н (9) | / | /
С=О С=О С=О С=О
| | | |
НО–С–Н + Н–С–ОН ¾® НО–С–Н + Н–С–ОН
| | | |
Н–С–ОН Н–С–ОН Н–С–ОН СН2О(Р)
| | | 3-фосфоглице-
СН2О(Р) СН2О(Р) Н–С–ОН риновый аль-
ксилулозо-5-фосфат эритрозо-4- | дегид
фосфат СН2О(Р)
фруктозо-6-фосфат
Образовавшийся в реакции 9 фруктозо-6-фосфат так же, как и в реакции 8, изомеризуется в глюкозо-6-фосфат, а цикл реакций, указанный выше, повторяется ещё раз до образования второй молекулы 3-фосфо-глицеринового альдегида. В этих реакциях происходит окисление еще трёх молекул глюкозо-6-фосфата, однако две из них регенерируются из фруктозо-6-фосфата. Одна из молекул 3-фосфоглицеринового альдегида под действием фермента триозофосфатизомеразы изомеризуется в фосфодиоксиацетон, который далее взаимодействует со второй молекулой 3-фосфо-глицеринового альдегида с образованием фруктозо-1,6-дифосфата. Эту реакцию катализирует фермент альдолаза:
|
|
| |
Полученный в данной реакции фруктозо-6-фосфат, как и в реакции 8, изомеризуется в глюкозо-6-фосфат, который вновь может включаться в реакции пентозофосфатного цикла.
Таким образом, в результате окисления шести молекул глюкозо-6-фосфата в реакциях пентозофосфатного цикла шесть раз происходит реакция декарбоксилирования с образованием шести молекул СО2, при этом 5 молекул глюкозо-6-фосфата регенерируются, поэтому фактически окислению подвергается одна молекула глюкозо-6-фосфата. При окислении каждой из шести молекул глюкозофосфата дважды прроисходят реакции дегидрирования и в конечном итоге в них синтезируются 12 молекул восстановленных динуклеотидов НАДФ×Н. Суммарное уравнение окисления глюкозо-6-фосфата в пентозофосфатном цикле можно записать в следующем виде:
С6Н11О6–(Р) + 12НАДФ+ + 7Н2О ¾® 6СО2 + 12НАДФ×Н + 12Н+ + Н3РО4
В реакциях пентозофосфатного цикла не происходят процессы субстратного фосфорилирования, а синтезируемые в этом цикле восстановленные динуклеотиды НАДФ×Н довольно медленно окисляются ферментами электронтранспортной цепи митохондрий, вследствиие чего пентозофосфатный цикл не является активным источником свободной энергии для синтеза АТФ. Однако восстановленные динуклеотиды НАДФ×Н обладают большим восстановительным потенциалом по сравнению с НАД×Н, в связи с чем активно используются в качестве восстановителей во многих окислительно-восстановительных реакциях, таких как синтез жирных кислот, восстановление нитритов, сульфатов, дисульфидных связей в пептидах и белках, превращение рибонуклеотидов в дезоксирибонуклеотиды и др.
В пентозофосфатном цикле осуществляется превращение гексоз в пентозы, которые могут выводиться из цикла и участвовать в различных биосинтетических процессах. Так, рибозо-5-фосфат является исходным соединением, с которого начинается синтез рибонуклеотидов, участвующих в образовании нуклеиновых кислот, макроэргических нуклеозидполифосфатов, а также коферментных группировок НАД, НАДФ, ФАД, ФМН, кофермента А. Рибулозо-5-фосфат, превращаясь в рибулозо-1,5-дифосфат, участвует в процессах темновой фиксации СО2, в которых также потребляются восстановленные динуклеотиды НАДФ×Н, синтезированные в пентозофосфатном цикле.
У растений и бактерий пентозофосфатный цикл является также важнейшим источником эритрозо-4-фосфата для синтеза шикимовой кислоты, из которой образуются многие ароматические и фенольные соединения.
В хлоропластах реакции пентозофосфатного цикла наиболее активно проходят в темноте, поддерживая в отсутствии света необходимый уровень НАДФ×Н для восстановительных процессов. А поддержание определённого уровня АТФ осуществляется за счет триозофосфатов, образующихся в пентозофосфатном цикле, которые превращаются в 3-фосфо-глицериновую кислоту, инициирующую реакции субстратного фосфорилирования.
Следует отметить, что в пентозофосфатном цикле синтезируются промежуточные метаболиты, которые могут включаться также в реакции гликолиза (фруктозо-6-фосфат, 3-фосфоглицериновый альдегид, фосфодиоксиацетон), поэтому между этими двумя дыхательными путями осуществляется постоянная связь. Взаимодействие между гликолизом и реакциями пентозофосфатного цикла осуществляется очень легко ещё и потому, что они локализованы в жидкой фазе цитоплазмы, в которой постоянно происходят процессы диффузии различных химических компонентов. В связи с этим очень трудно определить, какая из молекул гексозы и на каком этапе включается в реакции гликолиза или пентозофосфатного цикла. При благоприятных условиях развития растительных организмов 10 - 40% гексоз, окисляющихся при дыхании, подвергаются распаду через реакции пентозофосфатного цикла, а остальные преимущественно – в реакциях гликолиза и цикла Кребса.
В анаэробных условиях реакции цикла Кребса и пентозофосфатного цикла ингибируются и дыхание в основном осуществляется через гликолиз. Соотношение дыхательных реакций реализующихся в виде гликолиза и пентозофосфатного цикла, зависит от природы клеток и их функционального состояния. Активизация реакций пентозофосфатного цикла наблюдается при дефиците влаги, затемнении растений, недостатке элементов питания, поражении инфекцией, старении, интенсификации процессов, связанных с потреблением НАДФ×Н, рибозо-5-фосфата, рибулозо-5-фосфата, эритрозо-4-фосфата.