Биосинтез РНК
Рис. 6.2 Схема эксцизионной репарации
Репарация начинается с присоединения ДНК-N-гликозилазы к поврежденному основанию. Существует множество ДНК-N-гликозилаз, специфичных к разным модифицированным основаниям. Ферменты гидролитически расщепляют N-гликозидную связь между измененным основанием и дезоксирибозой, это приводит к образованию АП (апуринового-апиримидинового) сайта в цепи ДНК (первый этап). Репарация АП-сайта может происходить при участии только ДНК-инсертазы, которая присоединяет к дезоксирибозе основание в соответствии с правилом комплементарности. В этом случае нет необходимости разрезать цепь ДНК, вырезать неправильный нуклеотид и репарировать разрыв. При более сложных нарушениях структуры ДНК необходимо участие всего комплекса ферментов, участвующих в репарации (Рис. 6.2.): АП-эндонуклеаза распознает АП-сайт и разрезает возле него цепь ДНК (II этап). Как только в цепи возникает разрыв, в работу вступает АП-экзонуклеаза, которая удаляет фрагмент ДНК, содержащий ошибку (III этап). ДНК-полимераза b застраивает возникшую брешь по принципу комплементарности (IV этап). ДНК-лигаза соединяет 3¢-конец вновь синтезированного фрагмента с основной цепью и завершает репарацию повреждения (V этап).
Пострепликативнаярепарация включается в тех случаях, когда эксцизионная не справляется с устранением всех повреждений ДНК до её репликации. В этом случае воспроизведение поврежденных молекул приводит к появлению ДНК с однонитевыми пробелами, а нативная структура восстанавливается при рекомбинации.
Врожденные дефекты системы репарации являются причиной таких наследственных заболеваний, как пигментная ксеродерма, атаксия-телеангиэктазия, трихотиодистрофия, прогерия.
Транскрипция – первая стадия реализации генетической информации в клетке. В ходе этого процесса происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна последовательности одной из цепей ДНК. В отличие от репликации, при которой копируется вся хромосома, транскрипция протекает избирательно. Процесс управляется особыми регуляторными последовательностями, указывающими начало и конец участков ДНК, подлежащих транскрипции. Единицы процесса транскрипции несут информацию о структуре одного или нескольких белков. Участок ДНК, в котором заключена информация о структуре одного белка, называется структурным геном.Внутри этих участков существуют разрывы – интроны, которые не несут генетической информации, относящейся к синтезу белка, кодируемого данным геном. Кодирующие части гена называются экзонами.
Субстратами и одновременно источниками энергии для транскрипции являются рибонуклеозидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ). Процесс осуществляется ДНК-зависимой РНК-полимеразой, которая у большинства изученных организмов представляет собой комплекс 4 и более неидентичных субъединиц, выполняющих разные роли. В ядрах эукариот обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая 45 S пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК и 5 S рРНК.
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию (Рис. 6.3). Инициация начинается с активации промотора (знак начала транскрипции). Это происходит при участии особого белка – ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора – ТАТААА- (ТАТА-бокс). Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение конформации фермента и раскручивание спирали ДНК с образованием транскрипционной вилки, в которой матрица ДНК доступна для инициации синтеза цепи РНК. РНК-полимераза синтезирует небольшой олигонуклеотид. После этого к ней присоединяются факторы элонгации, значительно повышающие активность фермента и облегчающие расхождение цепей ДНК. РНК-полимераза перемещается вдоль молекулы ДНК и копирует одну из её цепей, последовательно присоединяя нуклеотиды в образующейся РНК в соответствии с принципом комплементарности. Синтез цепи РНК идет от 5¢- к 3¢-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой мРНК. По мере движения РНК-полимеразы растущая цепь РНК отходит от матрицы, а двойная спираль ДНК позади фермента восстанавливается. Когда РНК-полимераза достигает конца копируемого участка (терминатора), фермент и первичный транскрипт отделяются от матрицы. Этот этап происходит с участием факторов терминации.
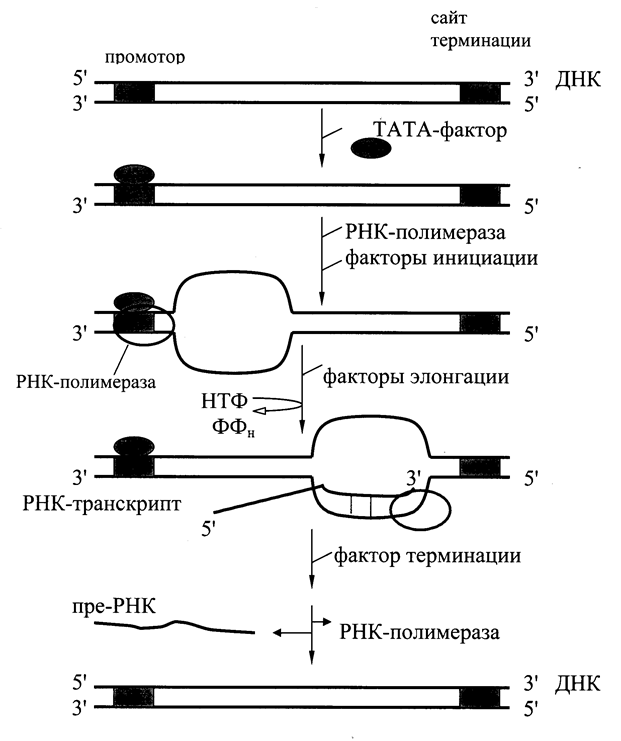
Рис. 6.3. Схема процесса транскрипции