АМИНОКИСЛОТЫ
ЛИПИДЫ
Липиды – нерастворимые в воде маслянистые или жирные вещества, которые могут быть экстрагированы из клеток неполярными растворителями. Это гетерогенная группа соединений, непосредственно или опосредовано связанных с жирными кислотами.
Биологические функции липидов:
1) источник энергии, который может длительно запасаться;
2) участие в образовании клеточных мембран;
3) источник жирорастворимых витаминов, сигнальных молекул и незамени-мых жирных кислот;
4) теплоизоляция;
5) неполярные липиды служат электроизоляторами, обеспечивая быстрое распространение волн деполяризации вдоль миелинизированных нервных волокон;
6) участие в образовании липопротеидов.
Жирные кислоты – структурные компоненты большинства липидов. Это длинноцепочечные органические кислоты, содержащие от 4 до 24 углеродных атомов, они содержат одну карбоксильную группу и длинный неполярный углеводородный «хвост». В клетках не встречаются в свободном состоянии, а только в ковалентно связанной форме. В состав природных жиров входят обычно жирные кислоты с четным числом атомов углерода, поскольку они синтезируются из двууглеродных единиц, образующих неразветвленную цепь углеродных атомов. Многие жирные кислоты имеют одну или несколько двойных связей – ненасыщенные жирные кислоты.
Наиболее важные жирные кислоты (после формулы приведено число атомов углерода, название, температура плавления):
 12, лауриновая, 44,2оС
12, лауриновая, 44,2оС
 14, миристиновая, 53,9оС
14, миристиновая, 53,9оС
 16, пальмитиновая, 63,1оС
16, пальмитиновая, 63,1оС
 18, стеариновая, 69,6оС
18, стеариновая, 69,6оС
 18, олеиновая, 13,5оС
18, олеиновая, 13,5оС
 18, линолевая, -5оС
18, линолевая, -5оС
 18, линоленовая, -11оС
18, линоленовая, -11оС

20, арахидоновая, -49,5оС
Общие свойства жирных кислот;
- почти все содержат четное число атомов углерода,
- насыщенные кислоты у животных и у растений встречаются в два раза чаще, чем ненасыщенные,
- насыщенные жирные кислоты не имеют жесткой линейной структуры, они обладают большой гибкостью и могут принимать разнообразные конформации,
- в большинстве жирных кислот имеющаяся двойная связь расположена между 9-м и 10-м атомами углерода (Δ9),
- дополнительные двойные связи обычно расположены между Δ9-двойной связью и метильным концом цепи,
- две двойные связи в жирных кислотах не бывают сопряженными, между ними всегда находится метиленовая группа,
- двойные связи практически всех природных жирных кислот находятся в цис-конформации, что приводит к сильному изгибу алифатической цепи и более жесткому строению,
- при температуре тела насыщенные жирные кислоты находятся в твердом воскообразном состоянии, а ненасыщенные жирные кислоты представляют собой жидкости,
- натриевые и калиевые мыла жирных кислот способны эмульгировать нерастворимые в воде масла и жиры, кальциевые и магниевые мыла жирных кислот растворяются очень плохо и не эмульгируют жиров.
В мембранных липидах бактерий встречаются необычные жирные кислоты и спирты. Многие из бактериальных штаммов, содержащих эти липиды (термофилы, ацидофилы и галлофилы), адаптированы к экстре-мальным условиям.
 изоразветвленные
изоразветвленные
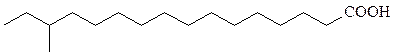 антеизоразветвленные
антеизоразветвленные
 циклопропансодержащие
циклопропансодержащие
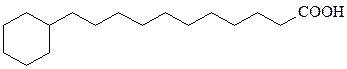 ω-циклогексилсодержащие
ω-циклогексилсодержащие
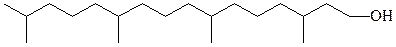 изопранильные
изопранильные
 циклопентанфитанильные
циклопентанфитанильные
Состав бактериальных липидов отличается большим разнообразием и спектр жирных кислот разных видов приобрел значение таксономического критерия для идентификации организмов.
У животных важными производными арахидоновой кислоты являются гистогормоны простагландины, тромбоксаны и лейкотриены, объединенные в группу эйкозаноидов и обладающие чрезвычайно широкой биологической активностью.
 простагландин Н2
простагландин Н2
Классификация липидов:
1. Триацилглицериды (жиры) – это эфиры спирта глицерола и трех молекул жирных кислот. Они составляют основной компонент жировых депо растительных и животных клеток. В мембранах не содержатся. Простые триацилглицериды содержат остатки одинаковых жирных кислот во всех трех положениях (тристеарин, трипальмитин, триолеин). Смешанные содержат разные жирные кислоты. По удельному весу легче воды, хорошо растворимы в хлороформе, бензоле, эфире. Гидролизуются при кипячении с кислотами или основаниями, либо под действием липазы. В клетках в обычных условиях самоокисление ненасыщенных жиров полностью заторможено благодаря наличию витамина Е, различных ферментов и аскорбиновой кислоты. В специализированных клетках соединительной ткани животных адипоцитах огромное количество триацилглицеридов может запасаться в виде жировых капель, заполняющих почти весь объем клетки. В форме гликогена организм может запасти энергию не более чем на сутки. Триацилглицериды могут запасать энергию на месяцы, так как они могут храниться в очень больших количествах в практически чистом, негидратированном виде и в расчете на единицу веса в них запасается в два раза больше энергии, чем в углеводах. Кроме того, триацилглицериды под кожей образуют теплоизоляционный слой, защищающий организм от действия очень низких температур.
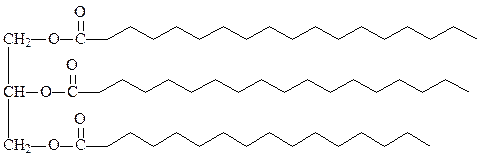 нейтральный жир
нейтральный жир
Для характеристики свойств жира используют следующие константы:
- кислотное число – количество мг КОН, необходимое для нейтрализации
свободных жирных кислот, содержащихся в 1 г жира;
- число омыления – количество мг КОН, необходимое для гидролиза
нейтральных липидов и нейтрализации всех жирных кислот,
содержащихся в 1 г жира;
- йодное число – количество граммов йода, связываемое с 100 г жира,
характеризует степень ненасыщенности данного жира.
2. Воска – это сложные эфиры, образуемые длинноцепочечными жирными кислотами и длинноцепочечными спиртами. У позвоночных животных секретируемые кожными железами воска выполняют функцию защитного покрытия, которое смазывает и смягчает кожу, а также предохраняет ее от воды. Восковым слоем покрыты волосы, шерсть, мех, перья животных, а также листья многих растений. Воска вырабатываются и ис-пользуются в очень больших количествах морскими организмами, особенно планктоном, у которого они служат основной формой накопления высококалорийного клеточного топлива.
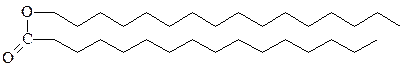
спермацет, получают из головного мозга кашалотов

пчелиный воск
3. Фосфоглицеролипиды – служат главными структурными компонентами мембран и никогда не запасаются в больших количествах. Обязательно содержат в своем составе многоатомный спирт глицерин, фосфорную кислоту и остатки жирных кислот.
Фосфоглицеролипиды по химическому строению можно еще разделить на несколько типов:
1) фосфолипиды – состоят из глицерина, двух остатков жирных кислот по 1-му и 2-му положению глицерина и остатка фосфорной кислоты, с которой связан остаток еще одного спирта (этаноламин, холин, серин, инозитол). Как правило, жирная кислота в 1-м положении насыщенная, а во 2-м – ненасыщенная.

фосфатидная кислота – исходное вещество для синтеза других фосфолипидов, в тканях содержится в незначительных количествах
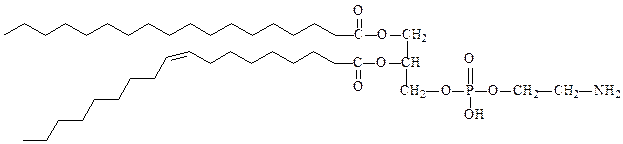 фосфатидилэтаноламин (кефалин)
фосфатидилэтаноламин (кефалин)
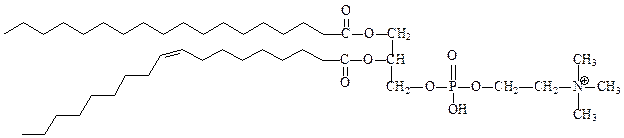 фосфатидилхолин (лецитин), его практически нет в бактериях
фосфатидилхолин (лецитин), его практически нет в бактериях
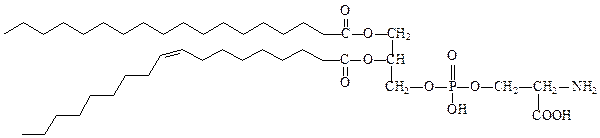 фосфатидилсерин
фосфатидилсерин

фосфатидилинозитол – предшественник двух важных вторичных мессенджеров (посредников) диацилглицерина и инозитол-1,4,5-трифосфата
2) плазмалогены – фосфоглицеролипиды, у которых одна из углеводо-родных цепей представляет собой простой виниловый эфир. Плазмалогены не встречаются в растениях. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
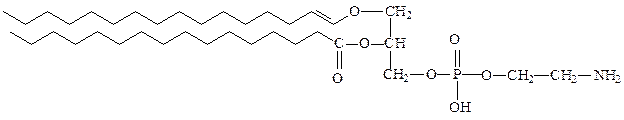 этаноламинплазмалоген
этаноламинплазмалоген
3) лизофосфолипиды – образуются из фосфолипидов при ферментатив-ном отщеплении одного из ацильных остатков. В змеином яде содержится фосфолипаза А2, которая образует лизофосфатиды, обладающие гемолитическим действием;
4) кардиолипины – фосфолипиды внутренних мембран бактерий и мито-хондрий, образуются при взаимодействии с глицерином двух остатков фосфатидной кислоты:
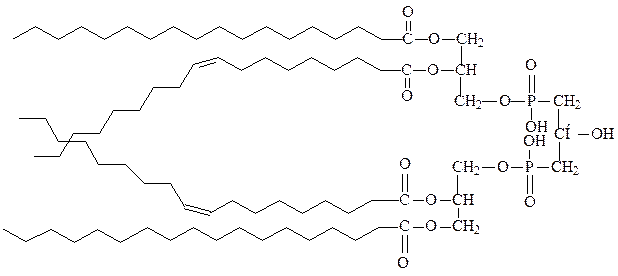 кардиолипин
кардиолипин
4. Фосфосфинголипиды – функции глицерина в них выполняет сфингозин – аминоспирт с длинной алифатической цепью. Не содержат глицерина. В большом количестве присутствуют в мембранах клеток нервной ткани и мозге. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Производные сфингозина, ацилированного по аминогруппе остатками жирных кислот, называются церамидами. Важнейший представитель этой группы – сфингомиелин (церамид-1-фосфохолин). Он присутствует в большинстве мембран животных клеток, особенно много его в миелиновых оболочках нервных клеток определенного типа.
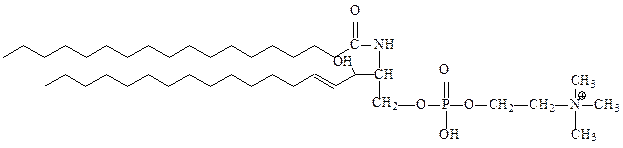 сфингомиелин
сфингомиелин
 сфингозин
сфингозин
5. Гликоглицеролипиды –липиды, у которых в положении 3 глицерола находится углевод, присоединенный с помощью гликозидной связи, фосфатной группы не содержат. Гликоглицеролипиды широко представлены в мембранах хлоропластов, а также в сине-зеленых водорослях и бактериях. Моногалактозилдиацилглицерол – наиболее распространенный в природе полярный липид, поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов:

моногалактозилдиацилглицерол
6. Гликосфинголипиды– построены из сфингозина, остатка жирной кислоты и олигосахарида. Содержатся во всех тканях, главным образом в наружном липидном слое плазматических мембран. В них отсутствует фосфатная группа и они не несут электрического заряда. Гликосфинголипиды можно разделить еще на два типа:
1) цереброзиды – более простые представители этой группы. Галактоцереброзиды содержатся главным образом в мембранах клеток мозга, тогда как глюкоцереброзиды присутствуют в мембранах других клеток. Цереброзиды, содержащие два, три или четыре остатка сахаров, локализуются в основном в наружном слое клеточных мембран.

галактоцереброзид
2) ганглиозиды – наиболее сложные гликосфинголипиды. Их очень крупные полярные головы образованы несколькими остатками сахаров. Для них характерно наличие в крайнем положении одного или нескольких остатков N-ацетилнейраминовой (сиаловой) кислоты, несущей при рН 7 отрицательный заряд. В сером веществе головного мозга ганглиозиды составляют около 6% мембранных липидов. Ганглиозиды – важные компоненты расположенных на поверхности клеточных мембран специфических рецепторных участков. Так они находятся в тех специфических участках нервных окончаний, где происходит связывание молекул нейромедиатора в процессе химической передачи импульса от одной нервной клетки к другой.
7. Изопреноиды – производные изопрена (активная форма – 5-изопенте-нилдифосфат), выполняющие самые разнообразные функции.
 изопрен
изопрен  5-изопентенилдифосфат
5-изопентенилдифосфат
Способность синтезировать специфические изопреноиды свойственна лишь некоторым видам животных и растений.
1) каучук – синтезируют несколько видов растений, в первую очередь гевея бразильская:
 фрагмент каучука
фрагмент каучука
2) жирорастворимые витамины А, D, Е, К (из-за структурного и функ-ционального сродства со стероидными гормонами витамин D сейчас относят к гормонам):
 витамин А
витамин А
 витамин Е
витамин Е
 витамин К
витамин К
3) гормоны роста животных – ретиноевая кислота у позвоночных и неоте-нины у насекомых:
 ретиноевая кислота
ретиноевая кислота
 неотенин
неотенин
Ретиноевая кислота является гормональным производным витамина А, стимулирует рост и дифференциацию клеток, неотенины – гормоны насекомых, стимулируют рост личинок и тормозят линьку, являются антагонистами экдизону;
4) растительные гормоны – абсцизовая кислота, является стрессовым фитогормоном, запускающим системный иммунный ответ растений, проявляющийся в устойчивости к самым различным патогенам:
 абсцизовая кислота
абсцизовая кислота
5) терпены – многочисленные душистые вещества и эфирные масла растений, обладающие бактерицидным и фунгицидным действием; соединения из двух изопреновых звеньев называются монотерпенами, из трех – сесквитерпенами, из шести – тритерпенами:
 камфора
камфора  тимол
тимол
6) стероиды – сложные жирорастворимые вещества, молекулы которых содержат в своей основе циклопентанпергидрофенантрен (по своей сути – тритерпен). Основной стерол в тканях животных – спирт холестерин (холестерол). Холестерин и его эфиры с длинноцепочечными жирными кислотами – важные компоненты липопротеинов плазмы, а также наружной клеточной мембраны. Из-за того, что четыре конденсированных кольца создают жесткую структуру, присутствие холестерина в мембра-нах регулирует текучесть мембран при экстремальных температурах. В растениях и микроорганизмах содержатся родственные соединения – эргостерин, стигмастерин и β-ситостерин.
 холестерин
холестерин
 эргостерин
эргостерин
 стигмастерин
стигмастерин
 β-ситостерин
β-ситостерин
Из холестерина в организме образуются желчные кислоты. Они обеспечивают растворимость холестерина в желчи и способствуют перевариванию липидов в кишечнике.
 холевая кислота
холевая кислота
Из холестерина образуются также стероидные гормоны – липофильные сигнальные молекулы, регулирующие обмен веществ, рост и репродук-цию. В организме человека основными являются шесть стероидных гормонов:


кортизол альдостерон


тестостерон эстрадиол


прогестерон кальцитриол
Кальцитриол – витамин D, обладающий гормональной активностью, он отличается от гормонов позвоночных, однако также построен на основе холестерина. Кольцо В раскрывается за счет светозависимой реакции.
Производным холестерина является гормон линьки насекомых, пауков и ракообразных – экдизон. Стероидные гормоны, выполняющие сигнальную функцию, встречаются также в растениях.
 экдизон
экдизон
7) липидные якоря, удерживающие молекулы белков или других соединений на мембране:
 убихинон
убихинон
Как мы видим, липиды не являются полимерами в буквальном смысле этого слова, однако как в метаболическом, так и в структурном отношении они близки к присутствующей в бактериях полиоксимасляной кислоте – важному запасному веществу. Этот сильно восстановленный полимер состоит исключительно из звеньев D-β-оксимасляной кислоты, соединенных сложноэфирной связью. Каждая цепь содержит около 1500 остатков. Структура представляет собой компактную правую спираль, около 90 таких цепей уложено с образованием тонкого слоя в бактериальных клетках.

поли-D-β-оксимасляная кислота
ЛЕКЦИЯ 4
Аминокислотами называются карбоновые кислоты, содержащие аминогруппу и карбоксильную группу. Природные аминокислоты являются 2-аминокарбоновыми кислотами, или α-аминокислотами, хотя существуют такие аминокислоты как β-аланин, таурин, γ-аминомасляная кислота. Обобщенная формула α-аминокислоты выглядит так:

У α-аминокислот при 2 атоме углерода имеются четыре разных заместителя, то есть все α-аминокислоты, кроме глицина, имеют асимметрический (хиральный) атом углерода и существуют в виде двух энантиомеров – L- и D-аминокислот. Природные аминокислоты относятся к L-ряду. D-аминокислоты встречаются в бактериях и пептидных антибиотиках.
Все аминокислоты в водных растворах могут существовать в виде биполярных ионов, причем их суммарный заряд зависит от рН среды. Величина рН, при которой суммарный заряд равен нулю, называется изоэлектрической точкой. В изоэлектрической точке аминокислота является цвиттер-ионом, то есть аминная группа у нее протонирована, а карбоксиль-ная – диссоциирована. В нейтральной области рН большинство аминокислот являются цвиттер-ионами:

Аминокислоты не поглощают свет в видимой области спектра, ароматические аминокислоты поглощают свет в УФ области спектра: триптофан и тирозин при 280 нм, фенилаланин – при 260 нм.
Для аминокислот характерны некоторые химические реакции, имеющие большое значение для лабораторной практики: цветная нингидриновая проба на α-аминогруппу, реакции, характерные для сульфгидрильных, фенольных и других групп радикалов аминокислот, ацелирование и образование оснований Шиффа по аминогруппам, этерификация по карбоксильным группам.
Биологическая роль аминокислот:
1) являются структурными элементами пептидов и белков, так называемые протеиногенные аминокислоты. В состав белков входят 20 аминокислот, которые кодируются генетическим кодом и включаются в белки в про-цессе трансляции, некоторые из них могут быть фосфорилированы, ацилированы или гидроксилированы;
2) могут быть структурными элементами других природных соединений – коферментов, желчных кислот, антибиотиков;
3) являются сигнальными молекулами. Некоторые из аминокислот являются нейромедиаторами или предшественниками нейромедиаторов, гормонов и гистогормонов;
4) являются важнейшими метаболитами, например, некоторые аминокислоты являются предшественниками алкалоидов растений, или служат донорами азота, или являются жизненно важными компонентами питания.
Классификация протеиногенных аминокислот основана на строении и на полярности боковых цепей:
1. Алифатические аминокислоты:
 глицин, гли, G, Gly
глицин, гли, G, Gly
 аланин, ала, А, Ala
аланин, ала, А, Ala
 валин, вал, V, Val*
валин, вал, V, Val*
 лейцин, лей, L, Leu*
лейцин, лей, L, Leu*
 изолейцин, иле, I, Ile*
изолейцин, иле, I, Ile*
Эти аминокислоты не содержат в боковой цепи гетероатомов, циклических группировок и характеризуется отчетливо выраженной низкой полярностью.
2. Серосодержащие аминокислоты:
 цистеин, цис, C, Cys
цистеин, цис, C, Cys
 метионин, мет, M, Met*
метионин, мет, M, Met*
Серосодержащие аминокислоты также малополярны, причем цистеин существует лишь в недиссоциированном состоянии. Благодаря образованию дисульфидных мостиков, цистеин выполняет важную функцию стабилизации пространственной структуры белков.
3. Ароматические аминокислоты:
 фенилаланин, фен, F, Phe*
фенилаланин, фен, F, Phe*
 тирозин, тир, Y, Tyr
тирозин, тир, Y, Tyr
 триптофан, три, W, Trp*
триптофан, три, W, Trp*
 гистидин, гис, H, His
гистидин, гис, H, His
Ароматические аминокислоты содержат мезомерные резонансно стабилизированные циклы. В этой группе только аминокислота фенилаланин проявляет низкую полярность, тирозин и триптофан характеризуются заметной, а гистидин – даже высокой полярностью. Гистидин может быть отнесен также к основным аминокислотам.
4. Нейтральные аминокислоты:
 серин, сер, S, Ser
серин, сер, S, Ser
 треонин, тре, T, Thr*
треонин, тре, T, Thr*
 аспарагин, асн, N, Asn
аспарагин, асн, N, Asn
 глутамин, глн,Q, Gln
глутамин, глн,Q, Gln
Нейтральные аминокислоты содержат гидроксильные или карбоксамидные группы. Хотя амидные группы неионогенны, молекулы аспарагина и глута-мина высоко полярны.
5. Кислые аминокислоты:
 аспарагиновая кислота (аспартат), асп, D, Asp
аспарагиновая кислота (аспартат), асп, D, Asp
 глутаминовая кислота (глутамат), глу, E, Glu
глутаминовая кислота (глутамат), глу, E, Glu
Карбоксильные группы боковых цепей кислых аминокислот полностью ионизированы во всем диапазоне физиологических значений рН.
6. Основные аминокислоты:
 лизин, лиз, K, Lys*
лизин, лиз, K, Lys*
 аргинин, арг, R, Arg
аргинин, арг, R, Arg
Боковые цепи основных аминокислот полностью протонированы в нейтраль-ной области рН. Сильно основной и очень полярной аминокислотой является аргинин, содержащий гуанидиновую группировку.
7. Иминокислота:
 пролин, про, P, Pro
пролин, про, P, Pro
Боковая цепь пролина состоит из пятичленного цикла, включающего α-углеродный атом и α-аминогруппу. Поэтому пролин, строго говоря, является не амино-, а иминокислотой. Атом азота в кольце является слабым основанией и не протонируется при физиологических значениях рН. Благодаря циклической структуре пролин вызывает изгибы полипептидной цепи, что очень существенно для структуры коллагена.
Некоторые из перечисленных аминокислот не могут синтезироваться в организме человека и должны поступать вместе с пищей. Это незаменимые аминокислоты отмечены звездочками.
Как было указано выше, протеиногенные аминокислоты являются предшественниками некоторых ценных биологически активных молекул.
Два биогенных амина β-аланин и цистеамин входят в состав кофермента А (коферменты – производные водорастворимых витаминов, образующие активный центр сложных ферментов). β-Аланин образуется путем декарбоксилирования аспарагиновой кислоты, а цистеамин путем декарбоксилирования цистеина:
 β-аланин
β-аланин  цистеамин
цистеамин
Остаток глутаминовой кислоты входит в состав другого кофермента – тетрагидрофолиевой кислоты, производного витамина Вс.
Другими биологически ценными молекулами являются конъюгаты желчных кислот с аминокислотой глицином. Эти конъюгаты являются более сильными кислотами, чем базовые, образуются в печени и присутствуют в желчи в виде солей.
 гликохолевая кислота
гликохолевая кислота
Протеиногенные аминокислоты являются предшественниками некоторых антибиотиков – биологически активных веществ, синтезируемых микроорганизмами и подавляющих размножение бактерий, вирусов и клеток. Наиболее известными из них являются пенициллины и цефалоспорины, составляющие группу β-лактамных антибиотиков и продуцирумые плесенью рода Penicillium. Для них характерно наличие в структуре реакционноспособного β-лактамного кольца, с помощью которого они ингибируют синтез клеточных стенок грамотрицательных микроорганизмов.
 общая формула пенициллинов
общая формула пенициллинов
Из аминокислот путем декарбоксилирования получаются биогенные амины – нейромедиаторы, гормоны и гистогормоны.
Аминокислоты глицин и глутамат сами по себе являются нейромедиаторами в центральной нервной системе.


дофамин (нейромедиатор) норадреналин (нейромедиатор)


адреналин (гормон) гистамин (медиатор и гистогормон)


серотонин (нейромедиатор и гистогормон) ГАМК (нейромедиатор)
 тироксин (гормон)
тироксин (гормон)
Производным аминокислоты триптофана является наиболее известный из встречающихся в природе ауксин – индолилуксусная кислота. Ауксины – это регуляторы роста растений, они стимулируют дифференцировку растущих тканей, рост камбия, корней, ускоряют рост плодов и опадение старых листьев, их антагонистами является абсцизовая кислота.
 индолилуксусная кислота
индолилуксусная кислота
Производными аминокислот также являются алкалоиды – природные азотсодержащие соединения основного характера, образующиеся в расте-ниях. Данные соединения являются исключительно активными физиологическими соединениями, широко используемыми в медицине. Примерами алкалоидов могут служить производное фенилаланина папаверин, изохинолиновый алкалоид мака снотворного (спазмолитик), и производное триптофана физостигмин, индольный алкалоид из калабар-ских бобов (антихолинэстеразный препарат):


папаверин физостигмин
Аминокислоты являются чрезвычайно популярными объектами биотехнологии. Существует множество вариантов химического синтеза аминокислот, однако в результате получаются рацематы аминокислот. Так как для пищевой промышленности и медицины пригодны только L-изомеры аминокислот, рацемические смеси необходимо разделять на энантиомеры, что представляет серьезную проблему. Поэтому более популярен биотехнологический подход: ферментативный синтез с помощью иммобилизированных ферментов и микробиологический синтез с помощью целых микробных клеток. В обоих последних случаях получаются чистые L-изомеры.
Аминокислоты используются как пищевые добавки и компоненты кормов. Глутаминовая кислота усиливает вкус мяса, валин и лейцин улучшают вкус хлебобулочных изделий, глицин и цистеин используются в качестве антиоксидантов при консервировании. D-триптофан может быть заменителем сахара, так как во много раз его слаще. Лизин добавляют в корм сельскохозяйственным животным, так как большинство растительных белков содержит малое количество незаменимой аминокислоты лизина.
Аминокислоты широко используются в медицинской практике. Это такие аминокислоты как метионин, гистидин, глутаминовая и аспарагиновая кислоты, глицин, цистеин, валин.
В последнее десятилетие аминокислоты начали добавлять в космети-ческие средства по уходу за кожей и волосами.
Химически модифицированные аминокислоты также широко используются в промышленности в качестве поверхностно-активных веществ в синтезе полимеров, при производстве моющих средств, эмульгаторов, добавок к топливу.
ЛЕКЦИЯ 5