Работа 58. Определение активности рибонуклеазы
РАЗДЕЛ 5. ОБМЕН НУКЛЕИНОВЫХ КИСЛОТ
Структурными единицами нуклеиновых кислот являются азотистые (пуриновые и пиримидиновые) основания, пентоза (рибоза и дезоксирибоза) и фосфорная кислота. Биосинтез пуриновых и пиримидиновых оснований осуществляется клетками большинства тканей. Пуриновое кольцо синтезируется из аминокислот глицина, глутамина, аспартата, производных тетрагидрофолата и СО2. Пиримидиновое кольцо синтезируется из карбамоилфосфата и аспартата. Нуклеотидные коферменты FAD+, NAD+, NADР+, CoA, являющиеся производными адениловой кислоты, синтезируются из ATP и соответствующих витаминов, а именно рибофлавина, никотиновой кислоты и пантотеновой кислоты. Предшественники ДНК - дезоксирибонуклеотиды образуются путем восстановления рибонуклеозиддифосфатов.
Конечными продуктами распада пуриновых оснований в процессе их обмена в организме являются мочевая кислота, аллантоин, аллантоиновая кислота, мочевина и глиоксалевая кислота., пиримидиновых оснований – карбаминовая кислота и b-аланин. Природа конечного азотсодержащего продукта, выводимого из организма, зависит от вида организма.
Работа 55. Спектрофотометрическое определение суммарного содержания нуклеиновых кислот
Для изучения природы и свойств нуклеиновых кислот необходимо выделение их из ткани в нативном, по возможности неизменном состоянии. Обычно этому препятствуют, главным образом, два обстоятельства: во-первых, крупные молекулы нуклеиновых кислот упакованы в структурах и прочно связаны с другими химическими компонентами клетки, в частности с белками; во-вторых, в клетках всегда есть активные нуклеазы, заключенные преимущественно в лизосомах и переходящие в свободное состояние при гомогенизации тканей. Таким образом, при выделении нуклеиновых кислот особое внимание необходимо уделять инактивации нуклеаз, полноте гомогенизации и очистке препарата от сопутствующих примесей (белки, полисахариды, полифосфаты и др.).
Методы количественного определения нуклеиновых кислот, основанные на спектрофотометрии в ультрафиолетовой области спектра, отличаются высокой чувствительностью и простотой проведения анализа. Необходимым этапом различных методов спектрофотометрического определения нуклеиновых кислот является их экстракция из биологического материала, сопряженная с гидролизом полинуклеотидов. В связи с этим нужно иметь в виду, что из исследуемого материала предварительно необходимо удалить свободные нуклеотиды.
В основе модификации спектрофотометрического метода определения суммарного содержания нуклеиновых кислот, разработанного А.С. Спириным, лежит экстракция их из биологического материала горячей хлорной кислотой с последующим определением поглощения экстрактов в ультрафиолетовой области спектра при 270 и 290 нм.
Исследуемый материал: ткани печени, сердца, скелетной мускулатуры крысы.
Реактивы:HClO4 – 0,2н и 0,5н растворы.
Оборудование: пипетки, центрифужные пробирки, пробки с воздушным холодильником, водяная баня, центрифуга, спектрофотометр, кюветы.
ХОД РАБОТЫ.Измельченные на холоде навески ткани печени, сердца, скелетной мышцы (100-200 мг) помещают в центрифужные пробирки и доливают в каждую по 5-10 мл охлажденного 0,2 н раствора хлорной кислоты. Содержимое пробирок тщательно перемешивают и осадок отделяют центрифугированием на холоде при 5000 об/мин в течение 10 мин. Надосадочную жидкость отбрасывают и осадок повторно отмывают хлорной кислотой. Такая предварительная обработка материала необходима для удаления кислоторастворимых нуклеотидов. После удаления супернатанта к осадку добавляют 5-10 мл 0,5 н раствора хлорной кислоты и, закрыв пробирки пробками с воздушным холодильником, нагревают их в кипящей водяной бане в течение 30 мин. Эта процедура обеспечивает количественную экстракцию нуклеиновых кислот из исследуемого материала и их кислотный гидролиз до растворимых фрагментов. Гидролизаты охлаждают и центрифугируют. Гидролизаты из одинаковых тканей объединяют и определяют поглощение на спектрофотометре при 270 и 290 нм против контрольного раствора – 0,5 н раствора хлорной кислоты. При необходимости гидролизаты разводят тем же раствором.
Рассчитывают содержание фосфора нуклеиновых кислот в 1 мл исследуемого раствора по формуле
С мкг Фн = (А270 – А 290) : 0,19,
где 0,19 – значение DА = (А270 – А290), которое имеет гидролизат нуклеиновых кислот, содержащий 1 мкг нуклеинового фосфора в 1 мл раствора. При дальнейших расчетах учитывают общий объем гидролизата и разведения. Для пересчета количества нуклеинового фосфора на количество нуклеиновых кислот пользуются средним пересчетным коэффициентом 10,3:
С мкг НК = С мкг Фн . 10,3.
Рекомендуется проводить дополнительное определение оптической плотности при 260 нм; оптическая плотность при 260 и 270 нм не должна различаться более чем на ±15%.
Работа 56. Определение температуры «плавления» водородных связей
Соответствующие пары азотистых оснований нативной ДНК упорядоченно связаны между собой водородными связями, температура плавления которых носит характер одномоментного кооперативного процесса. Поэтому при исследовании зависимости Ер от температуры в случае нативных образцов ДНК наблюдается ярко выраженный фазовый переход, т.е. в узком температурном интервале происходит резкий прирост значения Ер. В денатурированных образцах ДНК, где водородные связи не упорядочены, последние разрушаются постепенно, при разных температурах. Таким образом, исследование температурных зависимостей Ер является одним из тестов на нативность ДНК. Замечено, что значение температуры «плавления» ДНК коррелирует с содержанием в ее молекуле ГЦ-пар.
Исследуемый материал:препарат ДНК.
Реактивы: стандартный солевой раствор, содержащий хлорид натрия (0,15 моль) и цитрат натрия (0,015 моль) в 1 л (рН=7,0).
Оборудование: спектрофотометр с термостатированной камерой.
ХОД РАБОТЫ.Препарат ДНК растворяют (из расчета 10-20 мкг ДНК в 1 мл) в стандартном солевом растворе, разбавленном в 10 раз. Раствор помещают в кварцевую кювету (1 см) с герметической крышкой для предотвращения испарения. На спектрофотометре определяют экстинкции (Е260 нм) при разных температурах в промежутке от 25 до 95°С с интервалом не более 5°С и выдержкой при определенной температуре в течение 5-10 мин. Если наблюдается резкое изменение экстинкции при какой-либо температуре, это указывает на то, что исследуемая ДНК нативна; если рост экстинкции происходит постепенно, то ДНК частично денатурирована.
По результатам измерений строят кривую «плавления». Для этого по оси ординат откладывают отношения оптической плотности раствора при измеряемой температуре (t) к оптической плотности при 25°С, а по оси абсцисс – температуру. Точку плавления (tпл.) ДНК находят по кривой плавления. Она соответствует середине зоны подъема кривой относительной экстинкции.
Зависимость между точкой плавления и содержанием ГЦ-пар в ДНК определяют по формуле
сгц = (tпл. – 69,3) . 2,44,
где сгц – содержание ГЦ-пар в молярных процентах, а 69,3 и 2,44 – постоянные коэффициенты.
Средние данные получают из 3-4 параллельных опытов.
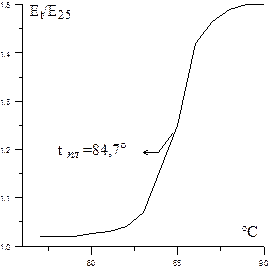
Кривая плавления ДНК, выделенной из микроорганизма Proteus vulgaris
Работа 57. Определение содержания мочевой кислоты
В организме человека мочевая кислота – конечный продукт расщепления пуринов, который выделяется с мочой. Исследование содержания мочевой кислоты в сыворотке крови используется в диагностических целях, поскольку увеличенное содержание мочевой кислоты в сыворотке крови наблюдается при некоторых заболеваниях, в частности подагре и синдроме Леша-Нейхана.
Принцип метода определения мочевой кислоты заключается в том, что мочевая кислота восстанавливает фосфорновольфрамовый реактив, а интенсивность образующейся окраски пропорциональна концентрации мочевой кислоты.
Исследуемый материал:сыворотка крови.
Реактивы: реактив Фолина (фосфорномолибденовый реактив), 20%-ный раствор ТХУ, насыщенный раствор Na2CO3, стандартный раствор мочевой кислоты.
Приготовление стандартного раствора мочевой кислоты. 100 мг мочевой кислоты вносят в колбу на 500 мл. Затем в нее приливают 25 мл 0,5%-ного раствора углекислого лития, 25 мл дистиллированной воды и после растворения мочевой кислоты доводят объем дистиллированной водой до метки, предварительно добавив для стойкости 12,5 мл формалина. Раствор стоек около месяца. Из основного (20%-ного стандартного раствора) готовят 2 %-ный рабочий раствор, разводя его в 10 раз. Рабочий раствор нестоек и может храниться лишь в течение 2-3 дней. 1 мл его содержит 0,2 мг мочевой кислоты.
Оборудование:центрифуга лабораторная, ФЭК, кюветы с длиной оптического пути 5 мм, центрифужные пробирки, стеклянные пробирки, пипетки градуированные, мерные колбы.
ХОД РАБОТЫ.В центрифужную пробирку вносят 1,5 мл сыворотки, 1,5 мл дистиллированной воды и 1,5 мл 20%-ной ТХУ. Содержимое пробирки тщательно перемешивают и через 30 минут центрифугируют при 3000 об/мин в течение 10 мин. К 1,5 мл супернатанта приливают 0,7 мл насыщенного раствора Na2CO3 и одну каплю реактива Фолина. Параллельно с опытной пробой обрабатывают стандартную. В пробирку помещают 0,5 мл стандартного рабочего раствора мочевой кислоты, 0,5 мл 20%-ного раствора ТХУ, 0,5 мл дистиллированной воды, 0,7 мл насыщенного раствора Na2CO3 и 1 каплю раствора Фолина. Через 10 мин. обе пробы колориметрируют на ФЭКе (зеленый светофильтр) в кювете с длиной оптического пути 5 мм против дистиллированной воды.
Расчетная формула
Сст . Еоп
Сх = ––––––––– ,
Ест
где
Сх – концентрация мочевой кислоты в сыворотке крови в мг%,
Сст – концентрация мочевой кислоты в стандартной пробе,
Еоп – экстинкция опытной пробы,
Ест – экстинкция стандартной пробы.
Норма содержания мочевой кислоты в сыворотке крови 2-3 мг%.
Нуклеиновые кислоты вовлекаются в деструктивный обмен при участии разнообразных нуклеаз. Существует несколько методов выявления их активности . Чаще всего пользуются измерением экстинкции при 260 нм до и после ферментативного гидролиза нуклеиновых кислот, реже - измерением содержания фосфора или пентоз. Активность нуклеаз определяют также по изменению вязкости раствора ДНК в процессе гидролиза и по величине гиперхромного эффекта.
Данный метод основан на том, что под влиянием рибонуклеазы (КФ 2.7.7.16) происходит расщепление фосфодиэфирных связей в молекуле РНК с образованием свободных кислых групп, которые обнаруживаются титрованием.
Исследуемый материал: разбавленная в 10 раз моча; кровь; 5%-ные водные гомогенаты тканей в количестве 0,5 мл.
Оборудование: холодильник, бюретка, пипетки на 1, 5, 10 мл, колбы конические на 50 мл.
Реактивы: раствор дрожжевой РНК (1%-ный) в ацетате натрия (0,1 М), NaOH (0,02 н.), фенолфталеин (0,5 %-ный спиртовой раствор).
ХОД РАБОТЫ.В две конические колбы вносят по 10 мл 1%-ного раствора дрожжевой РНК в 0,1 М растворе ацетата натрия. В одну колбу добавляют 0,2 мл 0,1%-ного раствора РНКазы, в другую – 0,2 мл воды. Содержимое колб перемешивают и смесь оставляют при 5°С на 1 ч. По истечении указанного времени из содержимого каждой колбы отбирают по 0,5 мл, добавляют по 2 капли фенолфталеина и титруют 0,02 н раствором гидроксида натрия до появления розового окрашивания. Оставшуюся смесь в колбах оставляют еще на 1 ч при 5°С. Затем из опыта и контроля берут пробы по 5 мл и оттитровывают их. Об активности РНКазы судят по разнице между количеством фосфатных групп в опытной и в контрольной пробах.
Если вместо раствора РНКазы используется слюна, кровь, моча или водные гомогенаты тканей, то вместо воды в контрольную пробу добавляют тот же объем биологической жидкости, что и в опыте, перемешивают, и немедленно фильтруют или центрифугируют и сразу же титруют 0,02 н раствором гидроксида натрия. Опытную пробу инкубируют 1-2 ч и обрабатывают затем подобным образом.