Практическое использование закона Харди-Вайнберга.
1.Одно из возможных применений закона Харди-Вайнберга состоит в том, что он позволяет рассчитать некоторые из частот генов и генотипов в случаях, когда не все генотипы могут быть определены вследствие доминантности аллелей. Например, зная количество особей с рецессивными признаками (аа) или соответственно q2, по формуле Харди-Вайнберга можно определить частоту рецессивных, а затем и доминантных генов в данной популяции.
Так, альбинизм у человека обусловлен довольно редким рецессивным геном. В популяциях Европы альбиносы рождаются с частотой 1:20 000 человек. Требуется определить частоту нормального доминантного аллеля и частоту нормально пигментированных людей, а также частоту гетерозиготных носителей гена альбинизма.
Если аллель нормальной пигментации обозначить А, а аллель альбинизма – а, то генотип альбиносов будет аа, а генотип нормально пигментированных людей – АА и Аа. Согласно закону Харди-Вайнберга, частота гомозигот (аа) равна q2; значит,
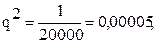 тогда
тогда  Так как p+q =1, то p=1-q, или
Так как p+q =1, то p=1-q, или
p=1-0,007=0,993, а частота доминантных гомозигот АА составит p2=(0,993)2=0,986.
Количество гетерозиготных носителей альбинизма 2pq =2´0,993´0,007=0,014. Тогда генотипическая структура европейской популяции такова: p2AA=0,98 595,
2pqАа =0,01400; q2аа =0,00005, или в абсолютных цифрах соответственно 19 719; 280 и 1.
2. Следующее важное применение закона Харди-Вайнберга состоит в том, что на основании расчетов можно выяснить одним или многими генами определяется тот или иной признак. Если частота распределения гомозигот и гетерозигот в популяции будет соответствовать расчетной, это значит, что данный признак или данное свойство определяется одним геном или одной парой аллелей.
Например, распределение людей по группе крови MN среди населения США следующие: 29,16% имеют группу крови MМ, 49,58% - MN и 21,26% - NN. Проверяем, соответствует ли это уравнению р2+2pq+q2 =1,т.е.
29,16ММ+ 49,58MN+ 21,26NN=100%. Частота генотипа NN,т.е. q2 =0,2126, а
 Частота р гена М=1-0,46=0,54, расчетная частота генотипа ММ р2=0,54´0,54=0,29, что согласуется с действительной частотой (0,2916 или 29,16%). Частота гетерозигот 2pq MN=2´0,54´0,46=0,49, что очень близко к 49,58% наблюдаемой частоты генотипа MN. Итак, р2+2pq+q2=0,29+0,49+0,21 или в процентах 29%MM+49%MN+21%NN, т.е. теоретические данные совпадают с фактическими. Это говорит о том, что группа крови MN действительно определяется одной парой кодоминантных генов.
Частота р гена М=1-0,46=0,54, расчетная частота генотипа ММ р2=0,54´0,54=0,29, что согласуется с действительной частотой (0,2916 или 29,16%). Частота гетерозигот 2pq MN=2´0,54´0,46=0,49, что очень близко к 49,58% наблюдаемой частоты генотипа MN. Итак, р2+2pq+q2=0,29+0,49+0,21 или в процентах 29%MM+49%MN+21%NN, т.е. теоретические данные совпадают с фактическими. Это говорит о том, что группа крови MN действительно определяется одной парой кодоминантных генов.
Этими же расчетами установлено, что одной парой аллельных генов определяется способность ощущать вкус фенилтиокарбамида, Хорея Гентингтона и ряд других заболеваний. Методами популяционной генетики установлено, что каждый сотый житель Европы несет в гетерозиготном состоянии ген амавротической идиотии, хотя заболевают 25 человек и миллиона. Даже в гетерозиготном состоянии мутантные гены часто в какой-то мере проявляются. Каждый 70-тый житель Европы гетерозиготен по гену альбинизма, хотя альбиносы встречаются с частотой 1:20 000.
Итак, зная правило Харди-Вайнберга, можно давать оценку генетического состава популяции человека. Но и это еще не все.
3.С помощью формулы Харди-Вайнберга определяют степень нарушения генетического равновесия природных популяций под действием внешних и внутренних факторов.
Допустим, что охотниками определенного материка за сезон добыто 16 000 шкурок лисиц, из них рыжих (АА) – 10 400, черно-бурых (Аа) – 4900 и черных (аа) – 700. Следует определить, находилась ли до промысла популяция в удовлетворительном состоянии и можно ли считать оправданным ее промышленное использование.
Определим, каким бы было соотношение этих трех генотипов лисиц в выборке, если бы популяция была идеальной. Частоту аллелей А и а рассчитываем по частоте генотипов (Табл.3).
Таблица 3
| Генотип и фенотип | Количество особей | Частота генотипов | Частота аллелей |
| АА (рыжие) Аа (черно-бурые) Аа (черные) Итого | 16 000 | 10400:16000=0,65 4900:16000=0,30 700:16000=0,05 1,00 | 
 1,00
1,00
|
Генетическая структура исследуемой популяции лисиц.
Теперь, воспользовавшись формулой Харди-Вайнберга, рассчитаем количество особей соответствующих генотипических классов в аналогичной выборке идеальной популяции:
р2АА = (рА)2´16 000 = 0,802´16 000 = 10 240
2pqAa = 2(pA)´(qa)´16 000 = 2´0,8´0,2´16 000 = 5 120
q2aa = (qa)2´16 000 = 0,22´16 000 = 640
Таким образом, аналогичная по численности выборка из идеальной популяции составляла бы из 10 240 рыжих, 5 120 черно-бурых и 640 черных лисиц, и приблизительно такое же количество шкурок было в добыче у охотников . С помощью метода х2 убеждаемся, что распределение особей между отдельными классами лисиц в данной реальной популяции достоверно не отличается от теоретически рассчитанного (р>0,050). Практически это означает, что генетическая структура популяции лисиц данного региона не нарушена и можно вести промысел в этой популяции. При других случаях можно было бы определить, какие факторы вызывали нарушение, и что ожидает популяцию в ближайшее и отдаленное время.
Одно интересное следствие из закона Харди-Вайнберга состоит в том, что редкие аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии. Рассмотрим приведенный выше пример с альбинизмом. Частота альбиносов (генотип аа) равна 0,00005, а частота гетерозигот Аа – 0,014, частота рецессивного аллеля а у гетерозигот составляет половину частоты гетерозигот, т. е.0,007.
Следовательно, при нужной частоте рецессивных аллелей в популяции они преимущественно находятся в гетерозиготном состоянии. Так, при частоте их 0,007 в гетерозиготе аллелей а примерно в 140 раз больше, чем в гомозиготе.
В общем случае, если частота рецессивного аллеля в популяции равна q, частота рецессивных аллелей в гетерозиготах составляет pq (половина от 2pq), а в гомозиготах – q2. Отношение первой частоты ко второй равно 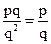 . Эта величина при малых значениях q приблизительно составляет
. Эта величина при малых значениях q приблизительно составляет 
Таким образом, чем ниже частоты аллеля, тем большая доля этого аллеля присутствует в популяции в гетерозиготном состоянии. Частота людей, страдающих алкаптонурией, равна
q2=0,000001, т.е. 1 на 1 млн., тогда как частота гетерозигот равна 2pq, т.е. около 0,002. Следовательно, число генов алкаптонурии в гетерозиготах примерно в 1000 раз больше, чем в гомозиготах.
Представим теперь, что некий диктатор, преследуя цель “улучшения расы” в соответствии с идеалом, решил элиминировать альбинизм (или алкаптонурию) из популяции. Поскольку гетерозиготы неотличимы от гомозигот по доминантному аллелю, его программа должна основываться на уничтожении или стерилизации рецессивных гомозигот. Это приведет лишь к весьма незначительному снижению частоты рецессивного аллеля в популяции, так как большинство аллелей альбинизма содержится в гетерозиготах, и, значит, не выявляется. Поэтому в следующем поколении частота альбиносов будет почти такой же, как и в предыдущем. Например, если исходная частота аллеля составляет 0,01, то для того, чтобы снизить ее до 0,001, потребуется 900 поколений (принято считать, что у человека смена поколений происходит каждые 25 лет), а чтобы достичь значения 0,0001 – 9 900 поколений. В общем случае число поколений t, необходимое для изменения частоты аллеля от q0 до q1, равно 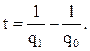
Равновесные частоты генов, сцепленных с полом.
Для сцепленных с полом генов равновесные частоты генотипов у самок (т.е. гомогаметного пола) совпадают с равновесными частотами аутосомных генов (т.е. р2для АА, 2pq для Аа и q2 для аа). Частоты генотипов гемизиготных самцов (т.е. гетерогаметного пола) совпадают с частотами аллелей: р для А и q для а. Самки с генотипом Аа получают одну гамету А от отца и вторую гамету А от матери; если частота аллеля А у самцов так же, как и у самок, равна р, то самки с генотипом АА будут появляться в потомстве с частотой р2. Аналогично частота самок с генотипом аа будет равна q2, а частота самок Аа – 2pq. Самцы, однако, всегда получают свою единственную Х-хромосому от матери. Поэтому частоты двух гемизиготных генотипов совпадают с частотами соответствующих аллелей у самок в предыдущем поколении.
Из этого следует, что фенотипы, определяемые рецессивными генами, у самцов встречаются чаще, чем у самок. Если частота сцепленного с полом рецессивного аллеля равна q, то частота определяемого им фенотипа будет равна q для самцов и q2 для самок. Отношение этих двух величин составляет q/q2 = 1/q. Чем меньше значение q, тем выше отношение частоты определяемого рецессивным геном фенотипа у самцов к его частоте у самок. Частота рецессивного сцепленного с полом аллеля, вызывающего дальтонизм у людей, составляет 0,08; следовательно, этот дефект встречается у мужчин в  раз чаще, чем у женщин. Частота рецессивного гена, определяющего наиболее распространенную форму гемофилии, равна 0,0001. В соответствии с законом Харди-Вайнберга, гемофилия у мужчин встречается в
раз чаще, чем у женщин. Частота рецессивного гена, определяющего наиболее распространенную форму гемофилии, равна 0,0001. В соответствии с законом Харди-Вайнберга, гемофилия у мужчин встречается в  раз чаще, чем у женщин (и при этом весьма редко у обоих полов: с частотой 1:10 000 у мужчин и 1:100 млн. у женщин).
раз чаще, чем у женщин (и при этом весьма редко у обоих полов: с частотой 1:10 000 у мужчин и 1:100 млн. у женщин).