Закон Харди-Вайнберга.
На первый взгляд, доминантное наследование признака, когда при встрече двух аллелей в зиготе один подавляет проявление другого, должно приводить к тому, что частота встречаемости доминантных генов от поколения к поколению будет увеличиваться.
Возникает вопрос: если короткопалость (брахидантилия) – аутосомно-доминантный признак, способный передаваться из поколения в поколение и концентрироваться в популяциях (согласно закону расщепления Менделя в соотношении фенотипов 3:1), то почему большинство людей не являются короткопалыми? Этого не происходит, потому что существует механизм поддержания частоты генов в популяции в равновесном состоянии. Закон равновесия генных концентраций панмиктической популяции был установлен английским математиком Г.Гарди (Hardy) и немецким врачом В.Вайнбергом (Weinberg) в 1908 году.
Основное утверждение закона Харди-Вайнберга состоит в том, что при наличии альтернативных и аллелей одинаковой жизнеспособности разных генотипов и при отсутствии элементарных эволюционных процессов (мутаций, отбора, миграций) исходное соотношение аллелей (независимо от их абсолютной частоты) сохраняется во всех последующих поколениях (популяционное равновесие). Этот закон утверждает также, что если скрещивание случайно (панмиксия), то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями.
Из закона Харди-Вайнберга вытекает следующий вывод: если частоты аллелей у самцов и самок исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигаются за одно поколение.
Если частоты аллелей у двух полов исходно различны, то для аутосомных локусов они становятся одинаковыми в следующем поколении, поскольку и самцы, и самки
получают половину своих генов от отца и половину – от матери. Таким образом, равновесные частоты генотипов достигаются в этом случае за два поколения. Однако в случае сцепленных с полом локусов равновесные частоты достигаются лишь постепенно (См. Сцепленные с полом гены). Следует отметить также, что такое случайное скрещивание.
Случайное скрещивание (панмиксия) происходит тогда, когда вероятность образования брачной пары между особями не зависит от их генетической конституции. Следовательно, в случайно скрещивающейся популяции частота спариваний носителей тех или иных генотипов пропорциональна доле, в которой эти генотипы представлены в популяции.
Закон Харди-Вайнберга в генетике аналогичен первому закону Ньютона в механике, который гасит, что любое тело сохраняет состояние покоя или равномерного прямолинейного движения до тех пор, пока действующие на него силы не изменят это состояние. Реальные тела всегда подвергаются воздействию внешних сил, но первый закон Ньютона служит отправной точкой для применения других законов механики. Закон Харди-Вайнберга гласит, что при отсутствии возмущающих процессов частоты генов не изменяются. Проверим это утверждение.
Создадим модельную популяцию диплоидных организмов, зададим закон их скрещивания, в котором отсутствует элемент случайности, и проследим, что произойдет с этой популяцией через несколько поколений. Представим также, что создаваемые нами организмы генетически просты: у каждого из них только по одному гену. Определим, что данный локус содержит один из двух аллелей, А и а, представленных с одинаковыми для самцов и самок частотами: p для А и q для а. Допустим, что самцы и самки скрещиваются случайным образом, или, что то же самое, гаметы самцов и самок образуют зиготы, встречаясь случайно. Тогда частота любого генотипа будет равна произведению частот соответствующих аллелей
(табл. ). Вероятность того, что определенная особь обладает генотипом АА, равна вероятности (p) получить аллель А от матери, умноженной на вероятность (р) получить аллель А от отца; т.е. р´р=р2. Аналогично вероятность того, что определенная особь обладает генотипом аа, равна q2.
Таблица 3.
Распределение двух аллелей при равновесном
состоянии популяции.
| Частоты гамет у самок | ||
| p(А) | q(a) | |
| p(A) | p2(AA) | pq(Aa) |
| q(a) | pq(Aa) | q2(aa) |
Генотип Аа может возникнуть двумя путями: организм получает аллель А от матери и аллель а от отца, или наоборот, аллель А от отца и аллель а от матери. Вероятность и того, и другого события равна pq, а, значит, суммарная вероятность
Возникновения генотипа Аа равна 2pq.
Теперь мы можем доказать справедливость трех утверждений, содержащихся в законе Харди-Вайнберга:
1. Частоты аллелей не изменяются от поколения к поколению. Это можно легко доказать. Частота аллеля А в потомстве в соответствии с табл.1равна сумме частоты генотипа АА и половины частоты генотипа Аа, т.е. равна р2+pq = p(p+q)= p (так как p+q=1). В том, что частота аллельных генов А и а в идеальной популяции со сменой поколений не изменяется, легко убедиться, подставив в бином (pA+qa)2 конкретные числа. Пусть у родительского поколения рА=0,4, qa=0,6. Тогда частота генотипов в F1 будет такая: (0,4А+0,6а)2 = 0,16АА + 0,48Аа + 0,36аа. Рассчитаем в F1 частоту генов А и а, исходя из частот генотипов:
pA  qa
qa 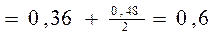
Таким образом, со сменой поколения частота аллельных генов в популяции не изменилась. Продолжая расчеты, можно убедиться, что соотношение аллелей А и а остается неизменным и в дальнейших поколениях.
2. Равновесные частоты генотипов задаются возведением в квадрат суммы частот аллелей и не изменяются от поколения к поколению. Так как частоты аллелей у потомства остаются такими же (p и q), какими были у родителей, то и частоты генотипов в следующем поколении также остаются постоянными и равными p2, 2pq и q2.
3. Равновесные частоты генотипов достигаются за одно поколение. Какими бы не были частоты генотипов в родительском поколении, частоты генотипов потомков будут p2, 2pq и q2, если частоты аллелей у самцов и самок одинаковы и равны p и q.
Формула Харди-Вайнберга применима при следующих условиях:
1) учитывается одна пара аутосомных генов; 2) спаривание особей и сочетание гамет в популяции совершается случайно; 3) обратные мутации настолько редки, что ими можно пренебречь; 4) достаточная многочисленность популяции; 5) особи разных генотипов имеют одинаковую жизнеспособность, плодовитость и не подвергаются элиминирующему отбору. К популяции самооплодотворяющихся организмов формула Харди-Вайнберга не применима.
В качестве примера соотношения Харди-Вайнберга используем данные о распределении белого населения США по группам крови системы M-N: ММ=1787, MN=3039, NN=1303, всего в выборке 6129 человек. На основании знания числа лиц с различными группами крови, мы можем рассчитать частоты аллелей. Частота аллеля LM равна сумме удвоенного числа индивидуумов с генотипом LM LM и числа индивидуумов с генотипом LM LN, деленной на общее число аллелей в выборке
(т.е. на удвоенное число обследованных лиц).
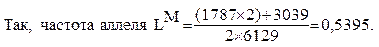
Точно так же рассчитывается частота аллеля LN; она равна 0,4605. Тогда отношение теоретически ожидаемых равновесных частот генотипов, рассчитанное в соответствии с законом Харди-Вайнберга составляет 0,2911 LMLM: 0,4968 LMLN: 0,2121 LNLN, что очень близко к реальному отношению генотипических частот, наблюдаемых в популяции (0,292 : 0,496 : 0,213).(Табл.)
return false">ссылка скрыта
Таблица
Частоты генотипов и аллелей в трех популяциях человека
по группам крови системы M-N.
| Популяция | Число обладателей группы крови | Всего | Частоты генотипов | Частоты аллелей | ||||||||||
| M | MN | N | LMLM | LMLN | LNLN | LM | LN | |||||||
| Аборигены Австралии | 0,030 | 0,296 | 0,674 | 0,178 | 0,822 nbsp; | 0,030 | 0,296 | 0,674 | 0,178 | 0,822 | ||||
| Индейцы Навахо | 0,845 | 0,144 | 0,011 | 0,917 | 0,083 | |||||||||
| Белые США | 0,292 | 0,496 | 0,213 | 0,539 | 0,461 |
Если в популяции для данного гена существуют три аллеля с частотами соответственно p+q+r =1, то частоты генотипов также соответствуют формуле биноминального распределения (p+q+r)2=p2+q2+r2+2pq+2pr+2qr =1. Если аллелей одного локуса n , то количество возможных генотипов в популяции вычисляют по