Синтез нуклеїнових кислот
ОБМІН НУКЛЕЇНОВИХ КИСЛОТ
Обмін нуклеїнових кислот. Розщеплення нуклеїнових кислот
Нуклеїнові кислоти потрапляють в організм з їжею у складі складних білків – нуклеопротеїдів. Внаслідок дії ферментів, які знаходяться у шлунковому та кишковому соках, нуклеопротеїди розщеплюються на прості білки та нуклеїнові кислоти. Прості білки далі розщеплюються до амінокислот, які всмоктуються. Утворені нуклеїнові кислоти розщеплюються під впливом ферментів нуклеаз, які поділяють на дві групи: ендонуклеази (розщеплюють зв’язки в середині молекул нуклеїнових кислот) й екзонуклеази (відщеплюють мононуклеоти від кінця полінуклеотидного ланцюга). Крім того, нуклеази поділяють на рибонуклеази, дезоксирибонуклеази та неспецифічні нуклеази.
З кишок у кров можуть всмоктуватись мононуклеотиди та нуклеозиди. Кров’ю вони переносяться до різних органів і тканин, де частина з них використовується для пластичних потреб, а інша частина розщеплюється далі та перетворюється на кінцеві продукти. Так, спочатку під впливом фосфатаз (нуклеотидаз) відщеплюється ортофосфатна кислота з утворенням відповідних нуклеозидів, а вони розщеплюються до азотистої основи та пентози. Ортофосфатна кислота використовується в організмі при утворенні кісткової тканини, для фосфорилювання органічних сполук та в багатьох інших процесах. Пентози беруть участь у реакціях, характерних для обміну вуглеводів.
Утворені пуринові та піримідинові основи розщеплюються до кінцевих продуктів. Сечова кислота – це кінцевий продукт обміну пуринових основ в організмі людини. При порушенні регуляції процесу синтезу сечової кислоти та зростанні її кількості в крові виникає гіперурикемія. Сечова кислота погано розчинна у воді, тому зростання її концентрації приводить до утворення кристалів і накопичення їх у суглобах під шкірою або у вигляді ниркових каменів.
Піримідинові основи (урацил, цитозин, тимін) поступово розщеплюються до амоніаку, карбон (IV) оксиду й інших сполук.
Синтез нуклеїнових кислот
Одночасно з розщепленням нуклеїнових кислот у тканинах організму постійно відбувається їх синтез з використанням енергії АТФ.
Процес біосинтезу ДНК називаєтсья реплікація (копіювання, самоподвоєння). Відбувається цей процес під час подвоєння хромосом, ДНК мітохондрій, хлоропластів еукаріот, а також у процесі поділу бактеріальних клітин і під час розмноження вірусів.
Реплікація ДНК відбувається за напівконсервативним механізмом (англ. conserve – зберігати) – у кожній новій синтезованій молекулі один ланцюг “старий”, одержаний з вихідної материнської ДНК, а другий – “новий”, синтезований (дочірній), тобто реплікація проходить так, що два ланцюги – вихідний і реплікований, сполучаючись між собою, утворюють наступне покоління молекул ДНК. Внаслідок реплікації з однієї вихідної молекули ДНК утворюються дві молекули – точні копії, репліки, абсолютно ідентичні одна одній і вихідній матричній молекулі ДНК. Напівконсервативний механізм синтезу ДНК забезпечує видоспецифічність передачі генетичної інформації від покоління до покоління.
Процес реплікації ДНК включає три етапи: ініціацію, елонгацію та термінацію.
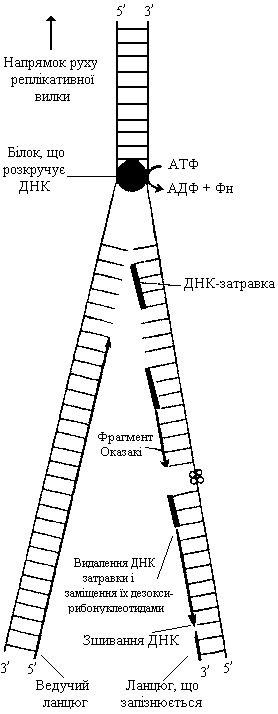
Ініціація полягає у формуванні реплікативної вилки та синтезі затравки. Транскрипція ДНК у ході поділу клітин починається з розділення двох ланцюгів, кожний з яких стає матрицею. Спеціальні ферменти та білки розплітають і розкручують подвійну спіраль ДНК з утворенням так званої реплікативної вилки – ділянки ДНК, у межах якої спіраль розкручена та розділена на окремі ланцюги.
|
| Реплікативна вилка |
У еукаріот утворюється від 10 до 100 тисяч стартових точок реплікації, тобто ділянок ДНК, з яких починається її розкручування. Якби у еукаріот була лише одна точка реплікації, то процес подвоєння ДНК тривав би декілька місяців (швидкість руху реплікативних вилок лише мікрометр за хвилину). Всі хромосоми в клітині повинні реплікуватися одночасно, й одночасно в клітині працюють багато тисяч вилок. Кожна з точок реплікації розміщується в окремих ділянках полінуклеотидного ланцюга, і реплікація їх проходить незалежно. Такі самостійні одиниці реплікації називаються репліконами. ДНК при утворенні реплікативної вилки розкручується не по всій довжині, а невеликими ділянками, що містять до 2 тисяч неспарених основ.
Молекула ДНК розкручується й утворюються дві одноланцюгові матриці. Оскільки в двоспіральній молекулі ДНК ланцюги антипаралельні – один з них має напрямок 5′→3′, а інший – 3′→5′, то після розкручування утворюються одноланцюгові антипаралельні матриці. Напрямок реплікації одного ланцюга співпадає з напрямком реплікативної вилки (5′→3′), його назвали ведучим, а ланцюг, реплікація якого протилежна напрямку реплікативної вилки (3′→5′), – тим, що запізнюється.
Після розкручування подвійної спіралі та утворення реплікативної вилки наступає друга стадія ініціації – синтез затравного ланцюга РНК на структурі ДНК-матриці. Затравка містить вільну 3′-ОН-групу, на якій може розпочатись синтез ДНК з використанням вільних нуклеозидтрифосфатів. Функцію затравки виконують невеликі фрагменти РНК (8-10 нуклеотидів), комплементарні ДНК-матриці. Синтез затравного полінуклеотидного ланцюга відбувається на ведучому ланцюгу. Потім фермент, що каталізує синтез затравки (праймосома) переміщується на ланцюг, що запізнюється, й синтезує затравку на ньому. По ланцюгу ДНК, що запізнюється, праймосома рухається до наступного місця початку синтезу в напрямку, протилежному напрямку синтезу ДНК, але однонапрямленому з реплікативною вилкою, синтезує затравку і знову переміщується вперед. Енергія для руху праймосоми забезпечується АТФ.
Елонгація, або подовження полінуклеотидного ланцюга, включає процеси реплікації ділянок материнської ДНК і сполучення синтезованих ділянок між собою. Реплікація відбувається за човниковим механізмом: спочатку реплікація відбувається в напрямку 5′→3′ під час просування вперед по одному ланцюгу, а потім у напрямку 5′→3′ продовжується в протилежному напрямку по другому полінуклеотидному ланцюгу матричної ДНК. Таким чином, синтез ланцюга, що запізнюється проходить не безперервно, а окремими фрагментами, які дістали назву фрагментів Оказакі. Утворюються вони внаслідок приєднання до 3′-ОН-групи затравок 1 000-2 000 дезоксирибонуклеотидних залишків. Послідовність нуклеотидів синтезованих фрагментів комплементарна нуклеотидній послідовності відповідної ділянки ланцюга матриці.
Дезоксирибонуклеотиди (дАТФ, дГТФ, дЦТФ, дУТФ) з’єднуються між собою з відщепленням пірофосфосфорної кислоти (Н4Р2О7). Так здійснюється поступовий ріст нового полінуклеотидного ланцюга, комплементарного матричному ланцюгу ДНК. Нарощування фрагментів Оказакі триває доти, поки не досягне місця, де розташована нова затравка, з якої розпочинається ріст нового фрагменту полінуклеотидного ланцюга. Далі затравки видаляються, а фрагменти Оказакі сполучаються між собою й утворюють дочірній ланцюг ДНК. Два новосинтезованих ланцюги, сполучаючись із своїми комплементарними матрицями, утворюють дві дочірні молекули ДНК, кожна з яких містить один материнський і один дочірній ланцюги.
Термінація – завершення синтезу ДНК відбувається після зустрічі двох реплікативних вилок, які рухаються назустріч одна одній. Процес реплікації проходить зі швидкістю 1 000-2 000 нуклеотидів за секунду у прокаріот і близько 100 нуклеотидів у еукаріот. В процесі реплікації на 1010 пар нуклеотидів трапляється лише одна помилка. Така висока точність є важливою умовою передачі спадкової інформації від одного покоління до іншого. Пошкоджені ділянки ДНК або помилково вбудовані нуклеотиди видаляються в результаті дії спеціальних ферментів нуклеаз.
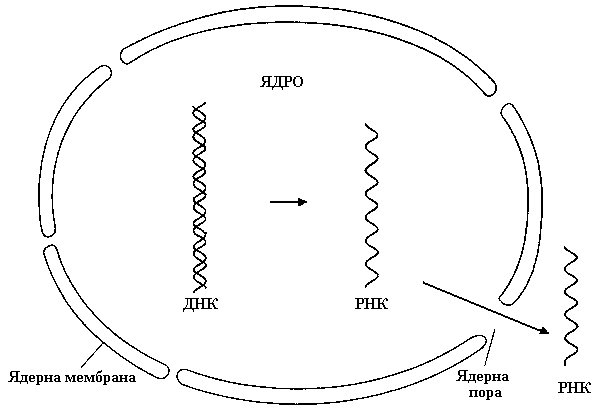
Процес біосинтезу РНК (і-, р-, т-) на матриці ДНК називають транскрипцією. Він здійснюється за принципом комплементарності, тобто нуклеотидна послідовність синтезованого ланцюга РНК комплементарна послідовності нуклеотидів одного з ланцюгів ДНК. Відмінність полягає лише в тому, що замість азотистої основи тиміну, у РНК включається урацил.
 Утворення та вихід з ядра молекули РНК
Утворення та вихід з ядра молекули РНК
|
Для синтезу РНК необхідна наявність усіх чотирьох видів нуклеотидів у формі трифосфатів (АТФ, ГТФ, УТФ, ЦТФ). У процесі їх сполучення відщеплюється пірофосфорна кислота (Н4Р2О7). Реакцію каталізує фермент РНК-полімераза (ДНК-залежна РНК-полімераза). Початок синтезу РНК не потребує наявності затравки. Спіраль ДНК частково розходиться, РНК-полімераза зв’язується з цією ділянкою ДНК і починається транскрипція РНК. Ріст (синтез) нуклеотидного ланцюга відбувається в напрямку 5′→3′. Швидкість транскрипції 40-50 нуклеотидів у секунду. Точність відтворення РНК у тисячі разів нижче за ДНК-реплікацію. Це пов’язано з тим, що транскрипція стосується лише однієї клітини, що не так важливо як реплікація, яка визначає майбутнє всього виду.
Синтез ДНК на матриці РНК. Деякі РНК-вмісні віруси за допомогою РНК-залежної ДНК-полімерази здатні синтезувати ДНК, комплементарну по відношенню до вірусної РНК. Вона вбудовується в геном еукаріотичної клітини, де може багато поколінь залишатися в прихованому стані. За певних умов (наприклад, дії канцерогенів) вірусні гени можуть активуватися, й здорові клітини перетворяться на ракові. Процес, що протікає у напрямку (РНК→ДНК) називається зворотна транскрипція. Каталізує процес фермент РНК-залежна ДНК-полімераза, або ревертаза (зворотна транскриптаза). Виявлення ферменту ревертази має важливе значення для ранньої діагностики онкологічних захворювань. Для функціонування ревертази необхідна затравка та матричний ланцюг РНК. Спочатку на одноланцюговій РНК-матриці за участю ферменту ревертази синтезується одноланцюгова ДНК. Внаслідок цього утворюється гібридна РНК-ДНК молекула, яка виконує роль матриці для синтезу комплементарного ланцюга ДНК. Цей етап каталізує фермент РНК-залежна ДНК-полімераза. На останньому етапі за участю ферменту РНК-ази від РНК відщеплюється молекула ДНК. Далі така ДНК включається в геном інфікованої клітини та зумовлює переродження її в ракову.