Развитие беспозвоночных
История беспозвоночных животных ведет свое начало от одноклеточных простейших организмов (Protozoa), которые ответвились от общего ствола с растениями в до-кембрийское время. Однако у первых примитивных представителей животного царства тело было мягким, поэтому они очень плохо сохранились в геологической летописи. Самые примитивные простейшие имеют общие признаки с одноклеточными зелеными водорослями. В современную эпоху простейшие (радиолярии и глобигерины) составляют значительную часть планктона морей il океанов и играют важную роль в океаническом осадкообразовании.
Все главные типы беспозвоночных животных возникли в докембрии и дали начало в высшей степени разнообразному миру водных и наземных форм в течение всего фанерозоя. Возникновение животных от их растительных предков, естественно, было связано с переходом от автотрофного способа питания к гетеротрофному. Этот переход, по-видимому, был постепенным, в течение длительного геологического времени. В докембрии происходило развитие различных водорослей, которые послужили основной питательной базой для морских животных.
В настоящее время царство животных, кроме хордовых, включает всех беспозвоночных, подразделяющихся на следующие главные типы:

Относительно находок самых древних представителей простейших, которые в дальнейшем послужили исходными формами для всех остальных животных, палеонтологические данные скудны и противоречивы. Одноклеточные животные организмы в древних докембрийских отложениях морфологически не отличаются от растительных микроорганизмов. Поэтому мы должны обратиться к известным промежуточным формам организмов между растениями и животными. К ним относятся одноклеточные жгутиконосцы (Flagellata) или, точнее, некоторые представители класса простейших. Но в пределах этого класса окрашенные жгутиковые объединяются в подкласс растительных жгутиконосцев (Phytomastigma), а бесцветные - в подкласс животных жгутиконосцев (Loomastigina). Жгутиконосцы занимают особое положение в мире животных и растений. Для них характерно наличие жгутиков - органов передвижения. Число их может быть различным - от единицы до нескольких штук, десятков и даже сотен. Среди жгутиконосцев встречаются организмы с разными типами обмена веществ, например с автотрофным и гетеротрофным, а некоторые представители сочетают в себе черты обоих способов питания.
В этом отношении особенно показателен отряд эвгленовых (см. рис. 2). Многочисленные виды рода Euglena широко распространены в современных пресноводных водоемах, и особенно в загрязненных водах. Многие виды эвглен изменяют характер питания в зависимости от условий окружающей среды. При хорошем освещении и при наличии растворенных минеральных веществ в воде эвглены обладают типичным автотрофньш способом питания. В их телах совершается фотосинтез с усвоением разных неорганических солей. В темноте в растворе, содержащем органические вещества, эвглены теряют хлорофилл, становятся бесцветными и усваивают из раствора готовые органические вещества.
Так у эвглен совершается переход от автотрофного питания к гетеротрофному. Часто развиваясь в загрязненных водах, в которых обильно растворено органическое вещество, эвглены сочетают оба способа питания - автотрофный и гетеротрофный. Если обесцвеченных в темных условиях эвглен вновь перенести на свет, то через короткий промежуток времени они снова зеленеют и переходят к автотрофному питанию.
Таким образом, у относительно примитивных одноклеточных организмов животный и растительный типы обмена веществ успешно сочетаются и свидетельствуют о тесной генетической связи обоих царств органического мира нашей планеты.
Простейшие животные по своему строению чрезвычайно разнообразны. Подавляющее их большинство имеет настолько малые размеры, что для их изучения необходимо пользоваться микроскопом. Тип простейших делится на несколько классов, среди которых можно отметить следующие: корненожки (Rhizopoda), споровики (Sporo-zoa) и инфузории (Infusoria). Фораминиферы из класса корненожек встречаются в ископаемом состоянии начиная с нижнего кембрия. Радиолярии представляют собой шаровидные формы с центральной капсулой и тонкими лучистыми ложноножками. Морские планктонные формы сохранились с докембрия. В частности, в черных кремневых сланцах нижних горизонтов серии Аделаида в Австралии найдены радиолярии, принадлежащие к грппе Spumellaria, возможно, присутствуют также представители Nosellaria.
Ископаемые, близкие к простейшим, обнаружены в Западной Африке, в пределах Нигерийского щита, в породах так называемой Бирримийской серии. В 1961 г. французские палеонтологи Р. Ховас и Р. Готкор нашли там ископаемые раковины (Protozoa) диаметром 35-800 мкм с кремневыми стенками. Исследователи предложили назвать их Birrimarpoldia, поскольку эти ископаемые относятся к самым ранним из известных в геологической летописи сро-раминиферам (радиологический возраст бирримия оценивается в 2 млрд лет).
В 1963 г. советский палеонтолог M. E. Зильбер описал формы радиолярий из свиты Имандра (Варзуга) в горизонте туфобрекчий и лав на Кольском полуострове.
Таким образом, все данные о находке простейших свидетельствуют об относительно раннем их появлении как представителей эукариот.
Одним из важнейших этапов в развитии живого вещества было возникновение многоклеточных организмов. Относительно происхождения многоклеточных были выдвинуты гипотезы такими выдающимися биологами, как Э. Геккель (1834-1919) и И. И. Мечников (1845-1916). Подавляющее большинство исследователей считают, что колониальность явилась необходимым этапом на пути к организации многоклеточных организмов. В принципе колониальность может легко возникнуть вследствие незаконченного бесполого размножения, когда клетки разделились, но не разошлись. Колонии такого типа часто образуются у бесцветных и зеленых жгутиконосцев, у инфузорий, солнечников, зеленых, синезеленых и диатомовых водорослей.
Вначале клетки, входящие в состав колонии, были одинаковыми. Но впоследствии началось их разделение в соответствии с выполняемыми функциями. Произошла дифференциация на клетки, воспринимающие пищу, определяющие подвижность и воспроизводительные. Различные факты из области анатомии и морфологии простейших, рассмотренные советским зоологом С. С. Шульманом [1974], свидетельствуют о том, что у многих групп простейших имеется определенная тенденция к переходу от одноклеточного к многоклеточному состоянию.
Одна из наиболее вероятных гипотез происхождения многоклеточности была выдвинута в 1935 г. видным советским зоологом В. А. Догелем (1882-1955). По его мнению, многоклеточность есть результат полимеризации. Установлено, что уровень организации живых существ, степень дифференциации и интеграции их частей теснейшим образом связаны с их размерами. Самые малые организмы представлены вирусами. Следующие по сложности - примитивные прокариоты заметно крупнее вирусов, но меньше одноклеточных эукариот. Наконец, многоклеточные эукариоты по своим размерам превосходят одноклеточных. Отсюда, естественно, вытекает: первые живые существа в истории биосферы 'были небольшими, что объяснялось их несовершенной организацией.
По мнению С. С. Шульмана, прогрессивная эволюция животных с момента возникновения клеточных организмов типа эукариот идет по схеме: организмы олигомерного строения -> организмы полимерного строения -> организмы олигомерного строения на новой основе. В данном случае автор усматривает проявление диалектического закона отрицания отрицания, при котором на последнем этапе имеет место не простое отрицание, а синтез.
На современном уровне знаний можно предполагать, что наиболее перспективными на первом этапе существования многоклеточных могли быть только плавающие примитивные животные, состоящие из недифференцированных клеток, активно добывающие пищу. Необходимость быстрого движения вызвала вначале дифференциацию клеток по способу питания и обмена веществ, движения, а затем все другие прогрессивные изменения, характерные для пол-царства многоклеточных животных. В связи со способами размножения и изменчивости, которые сохранились у многоклеточных в наследство от их предков, сами многоклеточные, по-видимому, даже повысили способность участвовать в дальнейших эволюционных преобразованиях, хотя бы по той причине, что усложнение организации открыло новые возможности изменчивости.
В качестве наиболее вероятных предков многоклеточных И. И. Мечников допускал бесцветных жгутиконосцев. Одной из существенных причин для возникновения первых многоклеточных было увеличение содержания свободного кислорода в биосфере. Другая важная предпосылка - появление эукариотических клеток.
Массовое развитие многоклеточных беспозвоночных животных отмечается в позднем докембрии, в венде. Выделение вендского периода было обосновано академиком Б. С. Соколовьш. В разных значительно удаленных друг от друга разрезах обнаружена фауна многочисленных беспозвоночных мягкотелых животных. Фауна этого типа первоначально была названа эдиакарской по названию местности Эдиакара в Южной Австралии. Она была обстоятельно описана М. Ф. Глесснером, изучившим свыше 1400 образцов с отпечатками и множество других остатков и следов животных. Для интервала времени 700-600 млн лет назад надежно установлены многочисленные мягкотелые организмы - морские беспозвоночные. Среди находок эдиакарской системы 70 % составляют кишечнополостные (Coelenterata). Из них ''A представляют собой меду-зоподобные плавающие формы либо неприкрепленные колониальные организмы вроде современных сифонофор Ve-lella. Типичные животные эдиакарской фауны показаны на рис. 16. Кишечнополостные представлены колониальными формами, прикрепленными ко дну моря наподобие современных морских перьев. Кроме этих животных, в составе эдиакарской фауны были морские черви, напоминавшие современных полихет, существа без панциря, похожие на членистоногих животных, и трехлучевые дискообразные организмы типа морских звезд. Все они оставили на поверхности дна следы и отпечатки своего тела. Некоторые организмы были довольно крупными. Например, обнаружена медуза почти 1 м в диаметре, морские перья более 1 м в длину. Листовидный морской червь Dickiыsonia длиной 1 м имел толщину менее 3 мм.
Венд Русской платформы с его сибирским эквивалентом юдомской серией хронологически совпадает с эдиакарской системой Южной Австралии. Благодаря исследованиям Б. С. Соколова установлено, что вендские отложения характеризуются большим разнообразием фауны бесскелетных, чем эдиакарская система. Вендская фауна СССР представлена десятками видов пелагических и бентосных родов книдарий, аннелид, членистоногих, своеобразных птеродинид (петалонамы), вероятно, моллюсков и пого-нофор с первыми следами хитинизации трубки. Среди остатков вендской фауны имеется ряд космополитических родов и видов. Вендский биологический комплекс более полиморфен, чем любой из предшествующих.

Наиболее существенные изменения в истории развития мира животных произошли на границе венда и кембрия. По меткому выражению В. В. Меннера и Н. А. Штрейса, это ''величайшая биостратиграфическая граница''. Она характеризовалась тем, что у морских животных стали формироваться твердые части тела: покровы, раковины, внутренний скелет. Уже раннекембрийская фауна коренным образом отличается от вендской. Правда, следует отметить, что в середине венда почти все бесскелетные беспозвоночные относились к фанерозойским типам и классам. Уже в кембрийский период особые сидячие формы бентоса - археоциаты получают широкое развитие. Ранее считалось, что археоциаты характерны только для кембрия. Однако исследователи стали сомневаться в том, что столь ярко выраженный тип животных смог возникнуть, просуществовать и полностью вымереть за столь короткий интервал геологического времени, тем более что столь близкие к археоциатам губки и некоторые кишечнополостные, зародившись в докембрии, прожили несравненно более длительный срок. Сейчас установлено, что археоциатоподобные формы обитали в позднем докембрии. Они найдены и описаны в геологических разрезах Евразии и Африки. Например, в чешском докембрии, в породах с нечетким стратиграфическим положением, А. Г. Волог-дип обнаружил остатки почти настоящих археоциат с двустенным скелетом.
Формирование твердых частей тела происходило у разных групп беспозвоночных в разное время. Так, у одних низкоорганизованных групп оно началось еще в середине венда, у других - в основном в кембрии, у более высокоорганизованных групп - моллюсков (наутилоидей, гастропод и двухстворчатых) и кораллов-ругоз - в ордовике.
Неорганические составные части твердых образований у современных организмов представлены преимущественно безводными карбонатами кальция (кальцит или арагонит), фосфатами кальция (апатит) и кремнеземом разных модификаций. Организмы позднего рифея и начала кембрия больше синтезировали апатиты, чем организмы более молодых геологических периодов. Карбонаты кальция, первоначально синтезировавшиеся лишь немногими эука-риотами, к середине кембрия стали обычными продуктами биоминерализации. Позже на основе биогенных карбонатов кальция возникли обширные биологические сообщества, из которых лучше всего известны коралловые рифы.
Кремнезем первоначально откладывался только губками. Впоследствии его стали широко использовать солнечники, радиолярии, диатомовые водоросли, а также некоторые зеленые растения.
Первоначальное усвоение кальция организмами было связано с их физиологическими функциями, и лишь впоследствии он стал осаждать фосфатные и карбонатные ионы, растворенные в морской воде, образуя твердые части тела. Однако до сих пор нет полной ясности в механизме построения твердых частей тела у организмов. Не разгаданы и причины этого, по-видимому, они носили экологический характер. Твердый скелет должен был служить опорой для организма и способствовать увеличению его размеров. Кроме того, твердый скелет, например, у морских губок позволил им занять большее пространство и соответственно поглощать большее количество пищи. У других животных скелет увеличил их прочность в качестве жесткой конструкции. Раковины у ранее мягкотелых животных предохраняли их от физических повреждений. Они стали служить защитой от хищников, которые появились позднее. В результате образования скелетов в кембрии возникли новые способы обитания в области морских мелководий. Губки приобрели возможность фильтровать бактерии, трилобиты - зарываться в донные отложения, моллюски - ползать по поверхности морского дна. Брахиоподы, мшанки и иглокожие с помощью скелетов смогли подниматься вертикально из воды, держась чуть-чуть над ее поверхностью, что позволило более эффективно фильтровать воду с микроорганизмами в целях получения пищи. Без твердых частей тела подобный образ жизни был бы невозможен или во всяком случае менее продуктивен. По мнению американского ученого Г. Хатчинсона, появление в начале фанерозоя скелетов, способных фоссилизироваться, отражает в основном возникновение хищничества, а до этого времени биосфера была в общем мирным царством, в котором защитные панцири не были нужны.
Филогенетические связи ряда важнейших типов беспозвоночных животных по данным палеонтологии и сравнительной анатомии представлены на рис. 17. Не вызывают сомнений достаточно обоснованные положения о происхождении губок и кишечнополостных от одноклеточных животных, низших червей - от кишечнополостных и не-мертин - от низших червей. Кажется также довольно вероятным и зарождение членистоногих от кольчатых червей (морских кольчатых-полихет).
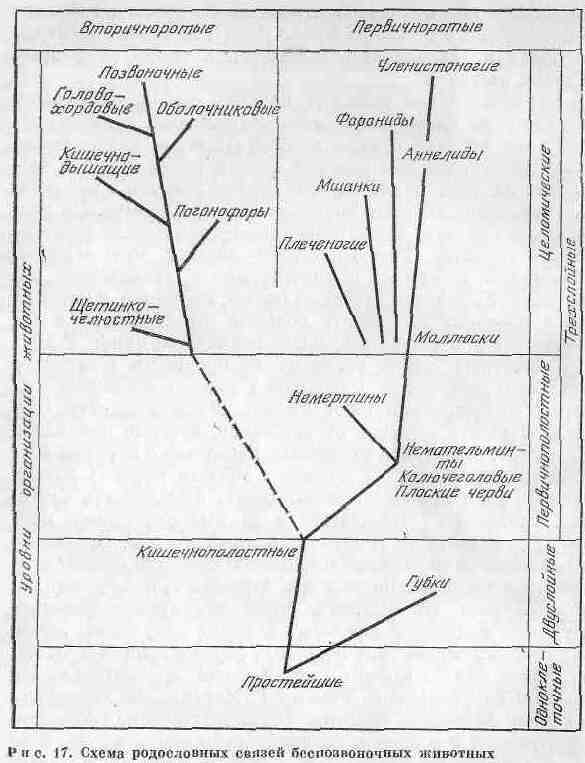
О возникновении других групп животных известно много меньше. Кроме того, следует учесть, что в основе рис. 17 лежит идея о монофилетическом происхождении животных, которую вполне можно принять для эволюции относительно молодых таксономических единиц. Однако для древних форм, корни которых уходят глубоко в до-кембрий, допустимо полифилетическое происхождение видов.
Рассмотрим кратко пути развития в фанерозое главных типов беспозвоночных животных.
Губки (Porifera или Spongia) являются преимущественно морскими животными, обитающими в пределах шельфа Все губки, (прикрепленные бентосные организмы) представляют собой примитивный тип многоклеточных животных. Их ткани, предназначенные для выполнения определенных функций, дифференцированы слабее, чем ткани других многоклеточных животных. Губки бывают кубковидными, шаровидными и произвольной формы с мелкими порами, через которые проникает вода с питательными веществами. Большое значение для систематики губок имеют формы иголок их скелета, называемая спикулами. По морфологии губок можно предположить, что они произошли от воротничковых и жгутиконосцев. Размножение губок бесполое и половым путем.
Начало свое губки ведут с позднего докембрия. А. 1. Вологдин и Н. А. Дроздова описали спикулы губок из гонамской свиты в Сибири, возраст которой 1500 млн лет.
Уже в кембрийских морях существовала довольно ьo-гатая фауна губок, представленная почти всеми классами и отрядами, дожившими до настоящего времени. Вымершие отряды восьмилучевых и разнолучевых губок встречаются в палеозойских отложениях. Наибольшего расцвета губки достигли в юрский и меловой периоды. Некоторые роды и даже виды меловых губок дожили до настоящего времени. В меловой период, вероятно, появились первые пресноводные губки.
Кишечнополостные. Тип кишечнополостных (Loelenterata) относится к лучистым беспозвоночным животным, для которых характерна радиально-осевая симметрия. Кишечнополостные, самые низкоорганизованные многоклеточные животные среди лучистых, обитают преимущественно в морской среде и значительно реже в пресноводных бассейнах. Группа кишечнополостных ведущих прикрепленный ко дну образ жизни, относится к полипам, а свободноплавающие мягкотелые - к медузам.
Полипы имеют цилиндрическую или бокаловидную форму Медузы по внешнему виду напоминают колокол или зонтик и плавают ртом вниз. Они бывают разных размеров, самые крупные медузы (Cyanea arctica) имеют зонтик диаметром до 2 м, щупальца растягиваются до 30 м Тело кишечнополостного организма чаще всего представляет собой мешочек, стенки которого состоят из наружного слоя, или экзодермы, и внутреннего, или эндодермы Внутри мешкоподобного тела находится пищеварительная полость с единственным отверстием наружу, называемым ротовым. Оно служит как для поглощения питательных веществ, так и для удаления отходов. Кишечнополостные обладают простым пищеварительным аппаратом. Непереваренная пища в виде остатков микропланктона и различные шлаки выводятся через ротовое отверстие. Кровеносная система у животных отсутствует. Нервная система состоит из отдельных нервных клеток, образующих неправильную сеть. Многие прикрепленные формы кишечнополостных имеют скелет, внутренний или наружный, органический или минеральный - известковый.
Большинство кишечнополостных раздельнополые организмы. Но наряду с половым размножением у них широко развито и бесполое, обычно посредством почкования. Иногда один и тот же вид кишечнополостных на разных стадиях жизненного цикла имеет строение то полипа, то медузы. По особенностям своего строения кшпечнополост-ные подразделяются на три класса.
Коралловые полипы образуют в настоящее время наиболее высокоорганизованную группу. Они представлены исключительно морскими одиночными и колониальными формами, ведущими прикрепленный образ жизни. У них отсутствует смена поколений и медузная стадия развития. Настоящие кораллы (Anthozoa - животные цветы) подразделяются на три подкласса: альционарии (восьмилучевые коралловые полипы), табуляты и зоонтарии.
Кишечнополостные, вероятно, возникли от свободно-плавающих многоклеточных организмов, которые уже имели клеточную организацию. Когда эти организмы начали вести прикрепленный образ жизни, появились Кишечнополостные гидроидные полипы, от них позднее произошли свободноплавающие медузы и коралловые полипы.
Первые находки кишечнополостных сделаны в разрезах верхнего докембрия. Наиболее древним считается отпечаток большой медузы, диаметром 18 см, в песчаном горизонте Нанкоуип серии Большого Каньона (штат Аризона, США). Отпечаток сохранился на поверхности тонкослоистого и тонкозернистого глинистого песчаника, сцементированного кремнеземом. По мнению американских геологов, серия Большого Каньона сопоставляется с серией Белт, возраст которой находится в пределах 1400-1000 млн лет.
Кишечнополостные наиболее древние из настоящих многоклеточных животных. Большое количество их (70%) обнаружено в эдиакарских отложениях Южной Австралии (см. рис. 16). В отложениях кембрийской системы найдены медузы и трубчатые кораллы - табуляты. В ордовике зарождаются более сложные формы - хететиды и четырехпучевые кораллы - ругозы. В конце пер-ми происходит массовое вымирание этих древних кораллов. В начале мезозоя появляются склерактинии и восьмилучевые кораллы - октокораллы. Эти две группы коралловых полипов продолжают существовать и в настоящее время.
Черви (Vermes). К ним ранее относили весьма разнообразных двусторонне-симметричных животных. В одном конце тела находится ротовое отверстие, а в противоположном - анальное. В настоящее время установлено, что обширная группа форм, объединяемая под названием Vermes, подразделяется на ряд типов. Среди них можно отметить тип плоских червей (Plathelnim-thes), круглых (Nemathelniinthes), немертин (Nemerti-ni) и кольчатых (Annelida). Поскольку черви в большинстве своем лишены твердых частей тела, они плохо сохраняются в ископаемом состоянии. Только в исключительно редких случаях черви доходят до нас в виде окаменелостей или оставляют после себя следы ползания в тонкозернистых породах.
Наиболее древние черви найдены в отложениях эдиа-кария-венда. Кольчатые черви по своему строению приближаются к типу членистоногих. Древние аннелиды, вероятно, были предками членистоногих.
Членистоногие (Arthropoda). Тип членистоногих берет свое начало в позднем докембрии, однако в фанерозое образовалось бесчисленное множество различных форм, резко превышающее число видов всех остальных типов животного мира. К настоящему времени на суше установилось господство двух типов животных - насекомых и позвоночных. Насекомых в составе морской и пресноводной фауны ничтожно мало. Они вторично приспособились к чуждой им водной стихии. Как отмечал В. И. Вернадский, если сравнить по массе людей и насекомых, то последние, по-видимому, преобладают среди форм наземной жизни. Это приводит к заключению, что насекомые являются исконными обитателями суши и их эволюционный путь совершался на поверхности континентов в обычной азот-кислородной атмосфере.
Поскольку, за редким исключением, все виды насекомых обитают на суше, естественно допустить, что, во-первых, для образования столь огромного числа видов потребовалось достаточно продолжительное геологическое время, захватывающее докембрий; во-вторых, насекомые явились первыми животными, населившими сушу. Следовательно, уже в докембрий (вероятно, в середине) должна была существовать благоприятная для этого среда. Главным же фактором было наличие свободного кислорода, необходимого для дыхания. Что касается питания, то здесь можно лишь предполагать. Например, что в прибрежных частях морских водоемов обитали водоросли, периодически попадавшие на сушу и доставлявшие корм наземным насекомым и их предкам. К настоящему времени в докембрийских отложениях не найдено никаких следов насекомых в ископаемом состоянии. Однако эти обстоятельства едва ли можно считать существенными, так как насекомые в ископаемом состоянии вообще встречаются значительно реже, чем другие представители тех же групп водных членистоногих. Это достаточно очевидно вытекает из наличия весьма нежного хитинового покрова, хрупкости их тел и образа жизни большинства насекомых. Даже в фанерозое в ископаемом состоянии насекомые встречаются крайне редко, Первичнобескрылые насекомые (из отряда Gollembola) впервые были найдены в отложениях верхнего девона. Но насекомые из отложений каменноугольной системы очень разнообразны: и приобретают хорошо развитые крылья и довольно крупные размеры, максимальные в истории мира насекомых. Эти данные, несомненно, говорят о том, что древнейшие насекомые появились гораздо раньше.
В геологических разрезах известны остатки также других классов членистоногих. К древним представителям членистоногих относятся трилобиты, мечехвосты, скорпионы и ракоскорпионы. Наибольшего размера достигали ракоскорпионы в силурийских морях, до 1 м в длину. Древнейшие скорпионы найдены в отложениях силурийской системы.
Моллюски (Mollusca). Тип моллюсков, или мягкотелых, представляет собой обособленную группу беспозвоночных животных, давших в фанерозое огромное число разнообразных форм, уступающее только членистоногим. Тело моллюсков состоит из головы, несегментированного туловища и ноги. Все моллюски относятся к симметрично-вторичнополостным животным. Для большинства из них характерна твердая минеральная раковина, часто покрывающая все тело животного. Хотя в настоящее время принято выделять 8-10 классов моллюсков, наибольшее значение в геологической истории имели только три: головоногие (Cephalopoda), брюхоногие (Gastropoda) и двустворчатые (Bivalvia). Все они сыграли выдающуюся роль в установлении стратиграфии фанерозоя. Большинство моллюсков водные животные, и лишь немногие представители класса брюхоногих вышли на сушу и приобрели легкие. Обособление моллюсков от других примитивных животных в процессе эволюции должно было произойти еще в докембрий.
Головоногие моллюски, получившие широкое развитие в раннем ордовике, вели кочевой образ жизни и питались живыми и мертвыми мелкими организмами. Позже некоторые из них, став плотоядными, перешли в разряд первых в истории животного мира хищников. Вероятно, в раннем палеозое существовали головоногие без скелетов, напоминавшие современных осьминогов и кальмаров. Головоногие моллюски были первыми животными, свободно плававшими в открытых водоемах на разной глубине. Они достигали высокой численности в морях ордовика. Длина отдельных животных составляла 10 м. В течение всего периода своего существования головоногие моллюски обитали в воде с постоянной соленостью. Ископаемые головоногие дали исключительное множество разнообразных форм. Однако в настоящее время сохранилось ограниченное число видов: осьминоги, кальмары, каракатицы и редкие наутилоидеи. В фанерозое особенно широко распространились наутилоидеи, аммониты и белемниты.
Ряд головоногих возникли в раннем кембрии. Уже в ордовике наутилоидеи обитали преимущественно на дне, в мелководных частях неритовой зоны. Первые формы наутилоидеи имели сильно сплющенную раковину и, по-видимому, ползали по морскому дну. В течение всего ордовика фауна наутилоидеи состояла преимущественно из форм с прямой раковиной. В то же время спирально свернутые раковины составляли примерно 15% всех головоногих. Но в дальнейшем соотношения резко меняются. В девоне наряду с прямыми и слабо изогнутыми формами существовало много спирально свернутых. Далее, в позднем палеозое, наутилоидеи с прямой раковиной становится значительно меньше, возрастает же число видов со спиральной раковиной. Спиральная ракови- на, гораздо более компактная и менее громоздкая, чем прямая, оказалась более удобной для свободного движения животного. Наутилоидеи со спирально скрученной раковиной быстро эволюционируют, приспосабливаясь к активному плаванию. Но плавали они относительно мало, невысоко над дном, на которое они могли опускаться из любого положения. Разнообразные и богатые формами раннепалеозойские наутилоидеи начинают постепенно вырождаться в девонский период, а с наступлением раннего карбона начинается их вымирание.
В силурийский период от спирально свернутых наутилоидеи происходят аммониты, а в девоне близкие к ним формы - гониатиты составляют уже большую группу морских организмов. Они более подвижны, чем их предки наутилоидеи. В течение девона, карбона, перми и всего мезозоя эволюционируют различные группы аммонитов. Их господство продолжалось до позднего мела и закончилось образованием гигантских форм, найденных в верхнемеловых отложениях. Так, один из верхнемеловых аммонитов в Вестфалии (ФРГ) достигал в диаметре 2,5 м. Аммониты существовали с начала девона до конца мела, и за это время сменилось около 1500 видов.
В палеогене произошло быстрое вымирание белемнитов. Причина этого еще недостаточно выяснена. Согласно наиболее распространенной точке зрения, массовое вымирание обширных групп головоногих моллюсков было связано с появлением в морях новых активных и высокоорганизованных противников. Ими могли быть костистые рыбы, обильно расплодившиеся в середине мелового периода, и морские млекопитающие - китообразные, которые заняли место морских рептилий мезозоя. В связи с появлением этих хищников исчезли не только аммониты, но и белемниты. Обе группы головоногих моллюсков, которые были подвижными в условиях мезозойских морей, оказались неуклюжими и неповоротливыми по сравнению с новыми хищниками. Поэтому в борьбе за существование уцелели самые сильные и самые проворные головоногие моллюски - типа осьминогов и кальмаров с сильно редуцированным известковым скелетом.
Изменения в строении раковин аммонитов, которые произошли на заключительных этапах их истории, позволяют предполагать, что в то время им пришлось вести борьбу с более подвижными хищниками, способными дробить раковины. Эту войну аммониты в конце концов проиграли.
Однако массовое вымирание аммонитов в конце мезозойской эры могло быть вызвано и другими факторами. Некоторые исследователи полагают, что вытеснение аммонитов костистыми рыбами не могло быть главной причиной их вымирания, поскольку аммонитами питались многочисленные хищные морские рептилии, в том числе гигантские мезозавры, ихтиозавры, плезиозавры и морские крокодилы. Однако при этом медлительные наутилоидеи выжили, а аммониты погибли. По мнению американского ученого К. Несиса, исчезновение подавляющего большинства известковых планктонных организмов было крупнейшим событием в мезозое. В результате погибли и гетеротрофные аммониты, которые ими питались.
Таким образом, общей причиной вымирания аммонитов была конкурентная борьба с костистыми рыбами, а конкретным поводом - внезапное вырождение известкового планктона - фораминифер и кокколитофорид. Но что было причиной внезапного исчезновения известкового планктона, неизвестно.
Остальные классы моллюсков - брюхоногие и двустворчатые - дожили до современной эпохи и дали большое разнообразие жизненных форм, в том числе обитающих в пресных водах. Самым распространенным оказался класс брюхоногих моллюсков с огромным количеством и ныне существующих видов. Представители класса брюхоногих моллюсков живут в морях, 'в солоноватых и пресноводных бассейнах, в реках и озерах, а отдельная группа их вышла на сушу и приснособилась к новому образу жизни. Множество ископаемых брюхоногих моллюсков найдено в отложениях фанерозоя, но максимального обилия и разнообразия они достигли в кайнозое.
Иглокожие (Echinodermata). Этот тип охватывает беспозвоночных животных, имеющих радиальную пятилучевую или реже двустороннюю (у некоторых раннепа-леозойских групп) симметрию мягкого тела. Они обладают внутренним известковым скелетом, построенным из пластинок, несущих иглы, откуда данный тип и получил название иглокожих. Для иглокожих характерна амбулак-ралъная, или водно-сосудистая, система, обеспечивающая их водой, кислородом и пищей. Все иглокожие - обитатели морей с нормальной или близкой к ней соленостью. Они размножаются половым путем. Все иглокожие делятся на два подтипа - Pelmatozoa 11 Elouterozoa.
К PeLmatozoa относятся все группы прикрепленных иглокожих бентоса. Они могут быть представлены и неприкрепленными формами, однако происходят все же от прикрепленных. К настоящему времени в морской фауне сохранился только один класс прикреплённых игаоко-жих - морские лилии. В геологическом прошлом существовало еще несколько классов иглокожих, ведущих прикрепленный образ жизни. К ним относятся морские пузыри (Cystoidea), текоидеи (Thecoidea):, карпоидеи (Саг-poidea) и морские бутоны (Blastoidea).
В эволюции иглокожих прикрепленные формы появились первыми. Довольно многочисленные представители иглокожих найдены в отложениях ордовика. Текоидеи встречаются от нижнего кембрия до нижнего карбона. Бластоидеи известны от ордовика до пермского периода включительно и в своем развитии дали формы, перешедшие от прикрепленного образа жизни к свободному, бентосному. Наиболее разнообразны формы морских лилий. Возможно, они произошли от цистоидей в кембрии или даже в докембрийское время.
В ордовике открыты остатки иглокожих, которые по своему анатомическому строению не укладываются ни в один из хорошо известных и общепризнанных классов. Сидячие формы иглокожих обитали на разных глубинах морского дна -от 20 до 5300 м. Большинство форм морских лилий живет в настоящее время на глубинах 4- 5 тыс. м, и лишь немногие - в мелких морях. В палеозое и мезозое морские лилии обитали преимущественно в мелководных прибрежных областях моря, некоторые переходили к свободному образу жизни. В различные эпохи возникали неприкреплепньге формы. Они известны в ордовике, девоне, карбоне и в отложениях меловой системы. Однако неприкрепленные формы оставались бентосными животными. Причина развития свободных форм связана с тем, что в определенных условиях способность передвигаться дает организму существенные преимущества перед формами, живущими на одном месте, там,, где животное, будучи личинкой, первоначально прикрепилось.
Подтип неприкрепленных иглокожих подразделяется на три класса: морских ежей (Ecliinoidea), морских звезд (Asterozoa) и голотурий (Holothurioidea). Свобод-ноживущие неприкрепленные иглокожие в отличие от прикрепленных составляют значительную часть современной морской фауны. Неприкреплонные формы иглокожих ведут свое начало с раннего палеозоя. В самом палеозое подвижные иглокожие дали большое количество разнообразных форм древних ежей, которые в конце палеозоя вымерли. Все свободноплаваюшие иглокожие являются в значительной мере хищниками, а также пожирателями трупов различных организмов.
К настоящему времени установлено, что тип иглокожих относится к наиболее высокоорганизованной группе беспозвоночных животных- вторичноротых. Первоначально -они были преимущественно двусимметричиымй, главным образом прикрепленными животными. Впоследствии из них выделились свободноплавающие неприкрепленпые формы.
Среди подвижных форм наиболее близки между собой морские звезды и офиуры. Первые известны с кем-брия, вторые - с силура. Допускают, что они выделились от ранпекембрийских примитивных эдриоастероидей (древних морских звезд). По всей вероятности, морские ежи имели родственные связи с последними. Однако наиболее древних ежей зоологи связывают с цистоидеями, а остальных - с эдраоастероидеями. Среди ныне живущих ежей наиболее примитивным считается отряд эластических ежей, большинство форм которых вымерло.
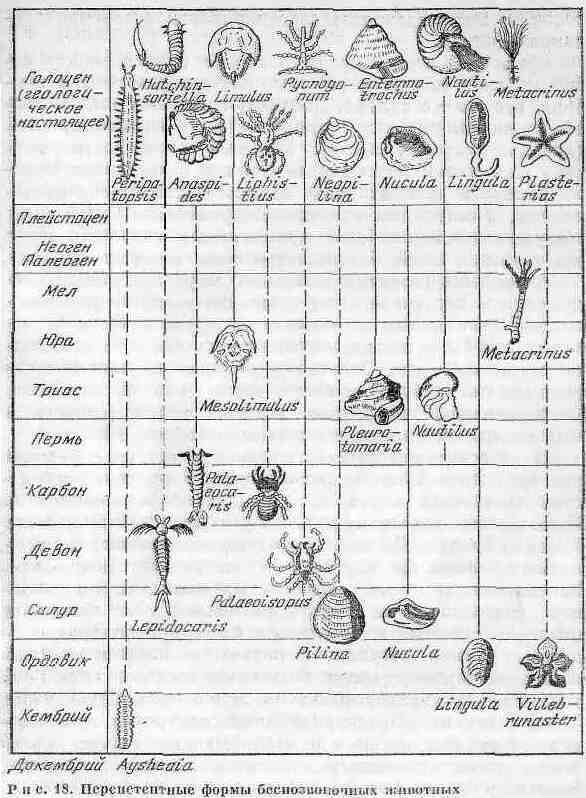
В процессе развития животного мира при общем прогрессивном поступательном движении имеются многочисленные ответвления определенных групп, эволюция которых как бы останавливается на десятки, а то и на сотни миллионов лет. Отдельные стороны перспстентности растений и микроорганизмов были уже рассмотрены. Беспозвоночные животные, которых можно отнести к персистентным формам, представлены на рис. 18.
В качестве классического примера обычно приводят род брахиопод Lingula, появившийся в ордовике и просуществовавший почти весь фанерозой без изменений. Столько же прожили роды двухстворчатых моллюсков Pilina a Nucula. Из головоногих моллюсков живым ископаемым является наутилус - ''жемчужный кораблик''. Представители этого рода - четырехжаберные с внешней, разделенной на камеры раковиной. Она защищает мягкие части тела животного, а благодаря наличию воздушных камер регулирует подводное плавание. Ныне живущий наутилус обитает премущественно в спокойной воде на глубинах от 40 до 700 м в юго-западной части Тихого океана. Предшественники наутилуса существовали от раннего пелеозоя до мезозоя включительно. К консервативным животным относится мечехвост Limulus, близкий к скорпионам и паукам.
Развитие позвоночных
Название ''позвоночные'' (Vertebrata) было введено в нау-.ку выдающимся французским натуралистом Ж. Б. Ламар-ком. Позвоночные как подтип животных возникли позже всех в биосфере. Их эволюция, начиная от первых примитивных предков .и кончая приматами и человеком, охватила фанерозой. Это наиболее высокоорганизованная, обширная и разнообразная группа животных из хордовых (Сhог-data). У представителей этого типа осевым скелетом является хорда - упругий эластичный стержень, простирающийся от хвостовой части тела животного до головной. В эмбриональном состоянии хорда присутствует у всех позвоночных, а у взрослых животных замещается чередованием хрящевых или костных позвонков. Центральная нервная система состоит из головного и спинного мозга. Головной имеет весьма сложное строение и помещается в черепной коробке (хрящевой или костной), которая есть у всех без исключения позвоночных. У большинства позвоночных животных две пары конечностей, которые в зависимости от среды обитания и характера движения меняют свои формы (плавники, ласты, ноги и крылья).
Позвоночные подразделяются на рыб (Pisces), земноводных (Amphibia), пресмыкающихся (Reptiоia), птиц (Aves) и млекопитающих (Mammalia).
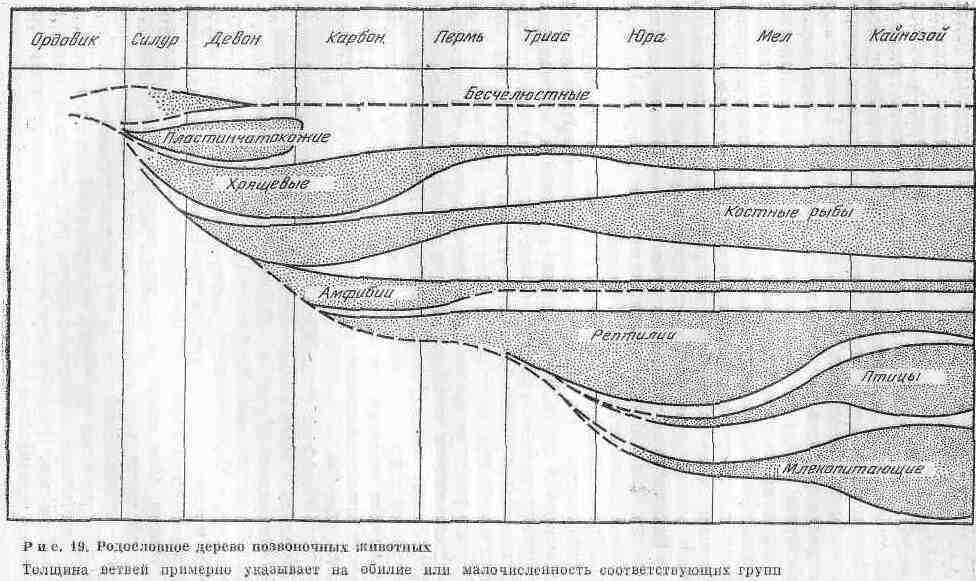
Происхождение позвоночных изучено лучше, чем других животных. По данным палеонтологии, сравнительной анатомии и эмбриологии, эволюция низших позвоночных первоначально совершалась в воде и привела к возникновению самых разнообразных круглоротых и рыб, которые приспособились к условиям обитания. Вероятнее всего, первые рыбы произошли от маленьких мягкотелых хордовых организмов, имевших хорду и жабры и живших за счет фильтрации частиц из морской воды. Переломным этапом в эволюции позвоночных был выход их на сушу. Предками наземных позвоночных были кистеперые рыбы, а древнейшими представителями сухопутных животных - земноводные (рис. 19).
Рыбы относятся к наиболее примитивным позвоночным. Из рыбообразных в самостоятельный надкласс выделяются бесчелюстные (Agnatha). Значительную их часть составляют круглоротые (Cyeloscomi), y которых челюсти отсутствуют. В современную эпоху они представлены миногами и миксинами, ведущими паразитический образ жизни. В ископаемом состоянии круглоротые известны с раннего ордовика. Они имели небольшие размеры, в передней головной части тела находился крупный плоский головной щит, состоящий из нескольких костяных пластинок. Глаза располагались по бокам, что связано было с придонным образом жнзни животных, когда они добывали пищу с морского дна. В процессе развития от позднего силура до раннего девона бесчелюстные дали большое разнообразие форм и приспособились к новым условиям жизни. Отдельные их виды приобрели; формы,: удобные, для плавания в открытом море. Они питались путем втягивания осадков трубкообразньш ртом, а также заглатывали и плавающий планктон с поверхности. Некоторые бесчелюстные рыбообразные были сходны с современными сомами, обитающими в пресноводных водоемах. Ископаемые остатки круглоротых свидетельствуют о том, что они рано приспособились к жизни в пресных водах. В раннем девоне бесчелюстные вели разнообразный образ жизни, но они должны были питаться очень мелкими организмами планктона либо тонкими осадками морского дна. Парадоксально то, что максимальное развитие примитивных круглоротых в девоне совпадает с развитием рыб с шарнирообразными челюстями. В конце девона они вытеснили круглоротых, у которых не было костного скелета.
Собственно рыбы (Pisces) имеют скелет, костный или хрящевой. У акуловых и осетровых сохранилась нерасчлененная хорда. Число позвонков у разных рыб резко различно-от 16 у рыбы луны до 400 у новозеландской рыбы ремня. Большинству рыб присуща удлиненная веретенообразная форма. Череп у костистых рыб состоит из большого числа костей; головной мозг хорошо развит, особенно у двоякодышащих рыб. Размеры рыб варьируют в широких пределах - от маленького бычка, длиной 1 см, обитающего на Филиппинских островах, до гигантской китовой акулы, достигающей 15 м в длину. Рыбы подразделяются на три класса: пластокожих (Placoderyes), хрящевых (Chondrichthyes), костных (Osteichthyes).
Первый класс представлен лишь ископаемыми палеозойскими формами; второй объединяет не только большое количество палеозойских форм, но и акул и скатов, живущих в настоящее время; третий охватывает наиболее прогрессивную группу, которая выделилась еще в палеозое, но достигла большого разнообразия в современную эпоху.
Пластинокожие объединяют наиболее примитивные формы челюстных, или настоящих, рыб. Тело их было покрыто пластинками, образующими своеобразный панцирь, который имел внешнее сходство с аналогичным панцирем круглоротых щиткообразпых форм. Поэтому ранее часть круглоротых и некоторых челюстных рыб объединяли в одну группу под названием ''панцирные''. Типичным представителем их являлся коккостеус около 50 см длиной. По общему очертанию тела это была обыкновенная рыба, но нижняя часть ее была плоской, что характерно для донных форм, и имела довольно длинный, заостренный к концу хвост.
Другой представитель пластинокожих - динихтис обладал мощным челюстным аппаратом с острыми выступами и краями, указывавшим на то, что рыба была хищником. Главное развитие пластинокожих пришлось на девонский период палеозоя. В прошлом существовало две группы плакодерм - артродиры и антиархи. Наиболее ранние артродиры были небольшими, несколько уплощенными рыбами, со слабыми челюстями. В течение девона они превратились в быстро плавающих хищников довольно крупных размеров. В ходе эволюции панцирь становился относительно короче и большая часть тела могла участвовать в движении. Совершенствовался челюстной механизм.
Антиархи были покрыты более обширным панцирем, почти по всей поверхности тела. Форма их была несколько плоской. Глаза быяи маленькими и находились в верхней части головного щита. Однако антиархи обладали сильными членистыми плавниками, внешне похожими на руки, приспособленные для перемещения по морскому дну. Ха-рактернои особенностью антиарх было наличие у них легких, позволяющих дышать воздухом.
Плакодермы не обнаруживают родственной связи с какой-либо группой современных рыб. Они вымерли в конце девона, уступив место более совершенным рыбам.
Хрящевые рыбы охватывают современных акул и скатов. Они имеют скелет примитивного строения, состоящий из хрящей. Наиболее ранние рыбы имели развитые наружные защитные образования, которые исчезли у более поздних форм. Хрящевые рыбы довольно древняя rpyuua. Первые из них появились в силуре. В дальнейшем число их возрастало, а в начале карбона развились многочисленные формы. Однако в конце палеозоя количество форм резко сократилось, а уже в настоящее время хрящевые рыбы составляют незначительную долю всей фауны рыб природных водоемов. Скаты оказались более современной ветвью акул.

Костные рыбы зародились в раннем девоне и первоначально обитали в пресноводных водоемах. В конце палеозоя они стали господствующей группой пресноводных позвоночных il заселили моря. Родословная и геохронология костных рыб представлены па рис. 20. В фауне современных водоемов большинство рыб относятся к классу костных. Они подразделяются на три подкласса: лучеперые с лучеобразными плавниками, двоякодышащие, кистеперые. Все они обнаружены в ископаемом состоянии в нижнем девоне и, возможно, происходят от акантодиев. У большинства современных костистых рыб имеется плавательный пузырь, регулирующий глубину плавания. Двоякодышащие рыбы обитали в пресных водах и, судя по строению тела, плавали всегда медленно. Они приспособились к жизни в периодически высыхающих реках и мелководных озерах. Легкие помогали им выжить в сухой сезон и в дальнейшем набраться сил в период дождей, когда в воде можно было дышать жабрами. В связи с этим двоякодышащие выжили в условиях, не переносимых другими рыбами. В настоящую эпоху существует лишь три вида двоякодышащих рыб, обитающих в реках Южной Африки, Австралии и Южной Америки, - своеобразные живые ископаемые.
Кистеперые рыбы (Crossopterygii) приспособились к активному образу жизни, давно заселили крупные реки и озера. Они представляют собой наиболее многочисленную группу, преобладающую в морях и теперь. Некоторые кистеперые (рипидисты), обитавшие в пресных водах, дали начало земноводным в позднем девоне. В этих условиях земноводные были более приспособленными хищниками, чем их предки, которых они вытеснили из мест обитания. Сохранившиеся рипидисты в карбоне и перми увеличили свои размеры и смогли жить в крупных реках и озерах.
В современную эпоху земноводные амфибии - самый малочисленный класс позвоночных. Они занимают особое место среди других позвоночных, поскольку являются первыми наиболее просто организованными четвероногими обитателями суши.
Земноводные дышат легкими, имеют два круга кровообращения, трехкамерное сердце и венозную и артериальную кровь. Температура тела зависит от температуры и влажности окружающей среды, что ограничивает возможность передвижения, ориентации и распространения на суше. Земноводные могут далеко уходить от природных водоемов. Однако в процессе размножения они неразрывно связаны с водой. Яйца амфибий как наземных животных лишены оболочек, предохраняющих их от высыхания, и в принципе не могут жить вне воды. В связи с водным способом рождения и обитания у земноводных развивается личинка, обитающая в воде. В дальнейшем личинка эволюционирует через ряд переходных форм (рыбоподобные, головастики и др.) и превращается в животное, обитающее на суше. В- настоящее время земноводные представлены тремя отрядами: хвостатых, безногих и бесхвостых.
Выходя в процессе эволюции на сушу, животные радикально изменяли конструкцию своего тела и главнейших органов. Морская вода, давшая жизнь позвоночным, содержала большое количество разнообразных растворенных питательных веществ, которые в условиях атмосферного воздуха отсутствуют. У первых живых организмов, выбравшихся на сушу, возникли проблемы, связанные с поисками пищи. Поскольку на многих географических широтах воздух, был недостаточно насыщен влагой, жизнь в условиях суши и атмосферного воздуха таила много опасностей, особенно для размножения, поскольку репродуктивные клетки весьма чувствительны к высыханию.
Соотношение свободного кислорода и углекислого газа в атмосферном воздухе отличается от водно-морского соотношения. Поэтому для сухопутных животных необходимы были механизмы, регулирующие процесс дыхания. Преломление световых лучей в воздушной среде ниже, чем в водной, что, в свою очередь, повлияло на строение глаз. Разная скорость распространения звука в воде и воздухе вызвала изменения слухового аппарата у животных. Наконец, резко изменились гравитационные условия существования организмов. В воздухе организмы весят больше, чем в воде. Следовательно, выход организмов из воды на сушу потребовал совершенствования физической опоры, что не могло не отразиться на развитии и прочности скелета. Таким образом, переход позвоночных животных на сушу и завоевание ее связаны с целым рядом изменений их тела в целях адаптация к новым условиям.
Первые представители земноводных имели крепкие кости, и их тела умеренных размеров были покрыты чешуей. По-видимому, их кожа практически не участвовала в процессе дыхания. Вместо нее для быстрого дыхания использовались легкие, изменившие объем с увеличением грудной клетки. У земноводных был длинный сильный хвост, увенчанный плавником, и четыре конечности, расположенные по бокам тела. По всей вероятности, тело и хвост были мощными средствами плавания, при этом в малой степени непосредственно участвовали сами конечности. Древние земноводные проводили много времени в воде. В водной среде первые амфибии с их острыми зубами, глазами, расположенными в верхней части головьг, и обтекаемой формой тела были похожи на небольших крокс-дилов.

По палеонтологическим данным, земноводные появились в раннем девоне. Родословная земноводных в масштабах геологического времени представлена па рис. 21. Предками земноводных были кистеперые рыбы, обладавшие легкими и парными -плавниками, из которых могли возникнуть пятипалые конечности. Такое заключение подтверждается поразительным сходством покровных костей черепа кистеперых с .костями черепа палеозойских земноводных. У земноводных животных и кистеперых рыб бьши как верхние, так и нижние ребра, которые отсутствовали у двоякодышащих рыб, ведущих в общем земноводный образ жизни. Двоякодышащие рыбы, обладавшие легкими, по целому ряду признаков отличались от земноводных, а значит, пе могли быть их непосредственными предками.
В девонский период происходило поднятие земной коры, обширные участки суши покрылись растительностью. Наибольшего разнообразия и численности земноводные достигли в каменноугольный и пермский периоды, когда, на обширных территориях господствовал влажный и теплый климат.
Ископаемые палеозойские земноводные относятся преимущественно к группе стегоцефалов, или панцироголовых.
Важной особенностью этих животных был сильный панцирь из кожных костей, покрывавший черепную коробку. В целом стегоцефалы отличались от современных земноводных рядом примитивных признаков, унаследованных от кистеперых рыб. К таким признакам, в частности, относится костный панцирь. Таз еще не соединялся с позвоночником, а плечевой пояс сохранял связь с черепом. Передние конечности были снабжены пятью пальцами.
Венцом развития древних земноводных был конец палеозойской эры, когда стегоцефалы достигали большой численности и разнообразия и еще не имели врагов среди более высокоорганизованных наземных позвоночных. Данные палеонтологии и анатомии земноводных позволили их подразделить на два подкласса - дугопозвонковых (Apsidospondyli) и тонкопозвонковых (Lepospondyli).
Первый подкласс охватывает надотряд лабиринтодонтов и надотряд прыгающих (Salicnlia), включающий многообразных современных амфибий.
Лабиринтодонты были найдены исключительно в ископаемом состоянии и представляли собой стегоцефалов, у которых имелся ''лабиринтодонтный'' тип зубов, полученный в наследство от их предков - кистеперых рыб. При поперечном срезе зубов обнаруживается необычно сложная картина разветвления эмалевых петель, в целом похожая на лабиринт. К группе лабиринтодонтов относились почти все крупные земноводные каменноугольного, пермского и триасового периодов. За время своего существования земноводные испытали существенные изменения. Более ранние формы имели умеренные размеры и рыбообразную форму тела; поздние укрупнились, тело их стало короче и толще и заканчивалось коротким толстым хвостом. Наиболее крупными представителями этих животных были мастодонзавры, обитавшие в триасе. Череп их достигал 1 м в длину. Внешним своим видом они походили на гигантских лягушек, питались главным образом рыбой. Мастодонзавры были преимущественно водными животными, редко покидавшими водную стихию.
Второй подкласс, тонкопозвоночных, охватывает три отряда стегоцефалов каменноугольного периода. Он был представлен мелкими, хорошо приспособленными к жизни в воде земноводными. Многие из них вторично утратили конечности и превратились в змеиноподобные формы. Предполагается, что из подкласса тонкопозвоночных амфибий в дальнейшем возникли современные отряды хвостатых и безногих земноводных.
В триасовый период одновременно существовали крупные земноводные и рептилии. Между ними происходила борьба за существование, которая закончилась победой рептилий, завоевавших в мезозое все области обитания поверхности планеты: воздух, сушу и водную стихию.
Пресмыкающиеся, или рептилии, в своей биологической организации имели явное преимущество перед земноводными. Они значительно лучше приспособлены к обитанию на суше. По существу, в истории развития позвоночных это были первые животные, которые размножались на суше яйцами, дышали только легкими, их кожу покрывали чешуйки или щитки. Развитие рептилий, даже обитающих в воде, не связано с водной средой. Волокнистая скорлуповатая оболочка яиц рептилий значительно препятствует их высыханию. По сравнению с амфибиями у рептилий более совершенная система кровообращения. Произошло дальнейшее разделение артериального и венозного токов крови. У большинства пресмыкающихся сердце трехкамерное, - однако желудочек разделен неполной перегородкой на две половины: правую - венозную и левую - артериальную. Несмотря-на прогрессивное развитие покровов, органов кровообращения и дыхания, пресмыкающиеся не Обеспечили себе теплокровность организма в целом, и температура их тела, подобно земноводным, зависит от температуры окружающей среды. Живущие в современную эпоху пресмыкающиеся подразделяются на четыре отряда: чешуйчатые, черепахи, крокодилы и клювоголовые. Пища пресмыкающихся довольно разнообразна, среди них встречаются насекомоядные, рыбоядные, хищники, растительноядные. В истории животного мира пресмыкающиеся прошли длительный путь развития. Первые находки рептилий известны в каменноугольных отложениях. Они относятся к отряду котилозавров из подкласса анапсид. Появившись в карбоне, они значительно развились в пермский период. Котилозавры были массивными животными и передвигались на толстых пятипалых конечностях. Размеры их колебались от нескольких десятков сантиметров до нескольких метров.
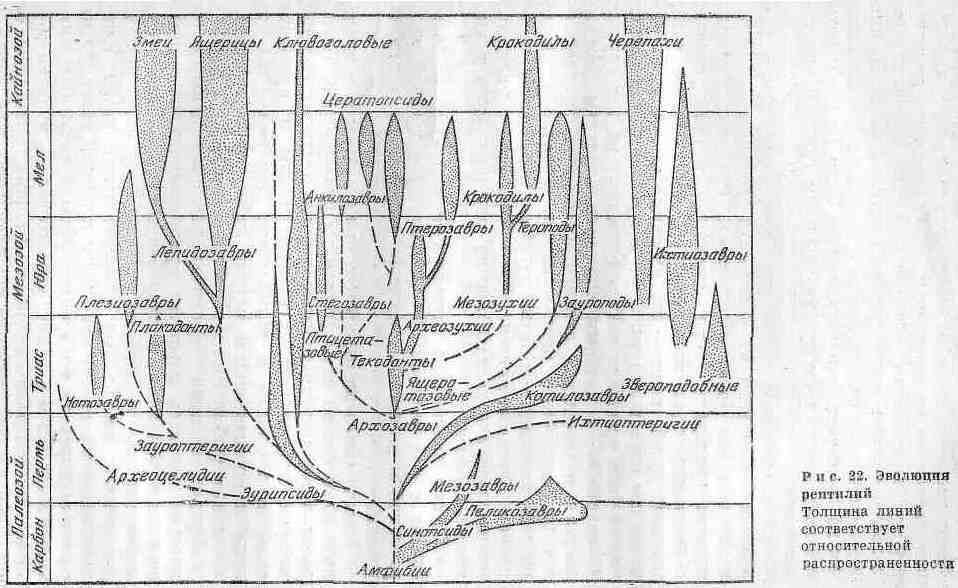
Родословные связи различных групп рептилий представлены на рир. 22. Наиболее примитивной формой из всех истаиаемых рептилий была сеймурия (Seymouria) - небольшое животное (от 0,5 м длиной), сохранившее ряд признаков земноводных. Практически у него отсутствовала шея, зубы имели примитивное строение, а кости черепа в деталях были сходны с черепной крышкой стегоцефалов. Детальное изучение сеймуроподобных рептилий и их остатков, найденных на территория СССР, позволило выделить группу батрехозавров, которые занимали промежуточное положение между земноводными и котилозаврами. По имеющимся данным палеонтологии и сравнительной анатомии можно заключить, что котилозавры были группой, которая дала начало всем другим основным группам пресмыкающихся.
От котилозавров произошли более подвижные формы рептилий. При этом их конечности удлинялись, скелет становился все более облегченным. Редукция черепного панциря проходила двумя основными путями: сначала образовалась одна височная яма, ограниченная снизу скуловой дугой, затем - две височные ямы: и соответственно две скуловые дуги. С учетом указанных признаков черепа всех пресмыкающихся разделяют па три группы: 1) анапсиды - с цельным черепным панцирем (котилозавры и черепахи) ; 2) синапсиды - с одной скуловой дугой (звероподобные, плезиозавры и, вероятно, ихтиозавры) ; 3) диапсиды - с двумя дугами, к ним относятся все прочие пресмыкающиеся. Последняя группа наиболее многочисленна и распадается на ряд подклассов и множество отрядов.
Группа анапсид по строению черепа была особенно близка к стегоцефалам. Из этой наиболее древней ветви рептилий к настоящему времени сохранились только черепахи.
Группа синапсид отделилась от котилозавров и привела к образованию ихтиоптеригий и синатозавров. Типичными представителями ихтиоптеригий были ихтиозавры, которые возвратились в водную стихию и приобрели сходство с рыбами и дельфинами. Они имели гладкую кожу и многочисленные острые зубы, приспособленные к питанию рыбой. Размеры их варьировали от 1 до 15 м.
Представителями синаптозавров были плезиозавры, которые являлись морскими пресмыкающимися, морфологически близкими к современным ластопогим. Они имели широкое утолщенное туловище, две пары мощных конечностей, измененных в плавательные ласты, длинную шею с маленькой головкой и короткий хвост. Размеры их варьировали в широких пределах: некоторые виды имели лишь 0,5 м в длину, но встречались и гиганты - 15 м. Они обладали сильными мыпщами, приводившими в движение ласты, которые служили лишь для гребли и не могли поддерживать корпус животного вне воды. Причины перехода от наземных форм синапсид к водным еще недостаточно выяснены.
Группа диапсид, объединяющая наиболее многочисленных и разнообразных рептилий, подразделяется на два подкласса - лепидозавров и архозавров. Еще в ранней перми выделились лепидозавры, известные как наиболее примитивные формы. От них произошли клювоголовые, чешуйчатые - ящерицы. Из клювоголовых в настоящее время известна гаттерия. В начале мелового периода от чешуйчатых отделились мезозавры - морские пресмыкающиеся, имевшие длинное (15 м) змеевидное тело и две пары конечностей в виде ластов. В конце мелового периода от ящериц отделилась новая ветвь - змеи.
От примитивного отряда эозухий, по всей вероятности, возникла большая ветвь - архозавров, распавшаяся впоследствии на три основные ветви: крокодилов, динозавров и крылатых ящеров.
Динозавры представляли собой наиболее многочисленную и разнообразную группу пресиыкающихся. Их размеры сильно варьировали - от мелких животных, величиной с кошку и меньше, до настоящих гигантов, достигавших почти 30 м в длину и весивших 40-50 т. На суше не было ничего подобного динозаврам. Масса наиболее крупных современных млекопитающих - слонов составляет лишь 1/5 массы крупного динозавра, а наиболее близкие родственники динозавров - крокодилы весят всего лишь около 1 т. Динозавры были легкими и массивными, подвижными и неуклюжими, хищными и травоядными, лишенными чешуи и покрытыми костным панцирем с различными выростами. Они разделялись главным образом по структуре таза на две большие группы - ящеротазовых и птицетазовых. У них были разные способы передвижения. В обеих группах динозавров встречались двуногие и четвероногие. Появились они в среднем триасе.
Ящеротазовые динозавры были представлены тероподами и завроподами. Тероподы относились в основном к плотоядным хищным животным, завроподы - преимущественно к травоядным. Ранние тероподы приспособились к быстрому бегу по ровной открытой местности. Некоторые тероподы по внешнему виду напоминали страусов. Они имели длинную шею и небольшую голову со слабыми челюстями, быстро бегали на длинных ногах и были всеядными. Наряду с быстро бегающими мелкими тероподами существовали и крупные животные, обладавшие огромной силой. Они были двуногими, опирались на мощные задние конечности и обладали малыми передними. К ним относились тираннозавр, цератозавр, аллозавр. Тираннозавр весил больше, чем самый крупный слон, достигал 10 м в длину и 6 м в высоту. Тираннозавры имели мощный череп с острыми зубами. Передние конечности были сильно уменьшенными и служили лишь для захвата пищи. Животные прыгали на задних конечностях, опираясь на хвост. Огромные тероподы, но всей вероятности, питались животными из группы ящеротазовых и птицетазовых.
В юрский и меловой периоды развивались завроподы. Это были самые крупные животные на Земле. Они обладали массивными скелетами, но у большинства форм задние конечности были длиннее и сильнее, чем передние. Это было связано с тем, что задние конечности должны были удерживать массивный таз, большую часть тяжелого позвоночника и длинный хвост. Передние же конечности были опорой только грудной клетке, сравнительно легкой шее и голове.
К крупным представителям ящеротазовых динозавров относились бронтозавр, диплодок, брахиозавр. Бронтозавр достигал в длину свыше 20 м, вес его приближался к 30 т. Диплодок был гораздо легче, но превосходил бронтозавра длиной (26 м). Неуклюжий брахиозавр был плотнее, его масса составляла около 50 т. Исходя из строения тела и массы этих динозавров, можно допустить, что все они, подобно современным бегемотам, большую часть времени проводили в воде. Они обладали слабыми зубами, пригодными для поедания лишь мягкой водной растительности, а у диплодока ноздри и глаза были сдвинуты кверху, что позволяло ему видеть и слышать, выставив из воды только часть головы.
Птицетазовые динозавры имели пояс задних конечностей, похожий на птичий. Они обладали средними размерами и разнообразными формами. Большинство из них передвигалось на четырех ногах. Животные были растительноядными, что отразилось на строении их зубов. имели, хорошо развитый панцирь, иногда покрытый разного рода наростами в виде рогов, шипов и т. д. Главное развитие птицетазовых динозавров приходилось на юрский и меловой периоды. Типичными представителями их были игуанодонты, стегозавры и трицератопсы.
Игуанодонты передвигались на задних ногах и достигали в высоту 5-9 м. Они были лишены панциря, первый палец передних конечностей представлял собой косой шип, могущий служить хорошим средством защиты от хищников. У стегозавров была крошечная голова, на спине располагался двойной ряд треугольных пластин, на хвосте несколько острых шипов. Длина стегозавров составляла 5-6 м.
В позднем мелу появились трицератопсы (церапсиды) - динозавры с рогоподобными выростами на черепе, служившими средством защиты от хищников. Трицератопс был похож на носорога. Большой рог размещался в конце морды, на носу, и пара рогов - над глазами. По размерам трицератопсы превосходили наиболее крупных носорогов, известных в настоящее время.
Птерозавры были первыми летающими позвоночными животными. Их передние конечности представляли собой настоящие крылья. К этой группе относится икарозавр, у которого ребра выдавались далеко в сторону. При полном выпрямлении ребер покрывающая их кожа легко могла превращаться в крыло. Летающие ящеры были не больше современной белки. По всей вероятности, они были не менее разнообразными, чем более крупные летающие формы, появившиеся в юрский и меловой периоды. Полагают, что икарозавры были лучше приспособлены к управлению несущими поверхностями и при этом у них сохранились развитые конечности, позволявшие хорошо ходить по земле, лазить и ощупывать различные предметы.
Настоящие птерозавры, или птеродактили, имели ряд общих черт с птицами: скрещенные грудные позвонки, большую грудину с килем, сложный крестец, полые кости, лишенную швов черепную коробку, большие глаза. Питались они преимущественно рыбой и жили в прибрежных скалах. У всех птерозавров тело было легким, с полыми костями, голова удлиненная и челюсти иногда со множеством зубов. Птерозавры были разными по величине - от размера воробья до гигантов, размах крыльев которых достигал 7 м. Птерозавры были представлены разнообразными формами. Среди них примитивная группа рамфоринхов, имевших длинный хвост для регулирования полета, и более высокоорганизованная группа собственно птеродактилей с зачаточным хвостом. Гигантские летающие ящеры - птеранодоны были самыми крупными летающими животными на Земле. При размахе крыльев до 7 м птеранодон весил 16 кг. Большая площадь крыла и легкая конструкция тела указывают, что птеранодоны преимущественно планировали и могли летать, взмахивая крыльями лишь в крайне необходимых случаях.
Из пермских котилозавров выделилась особая группа синапсид с развитыми челюстями, мощной мускулатурой и прогрессивной дифференциацией зубной системы - разнозубостью. Это сближало их с высшим классом позвоночных животных - млекопитающими.
В конце мелового периода произошло массовое вымирание мезозойских рептилий. Для объяснения этого события были предложены различные гипотезы, в том числе указывались космические факторы. Пожалуй, здесь нет необходимости заниматься критическим обзором всех этих представлений. Однако на одной вероятной причине следует остановиться. Большинство пресмыкающихся в течение всей мезозойской эры приобрели относительно высокую специализацию в разных средах обитания, которые были в общем довольно стабильными. В конце мезозоя орогенные процессы значительно усилились (эпоха ларамийского орогенеза), что отрицательно сказалось на условиях обитания рептилий. Дополнительно большую роль сыграла борьба за существование с другими, более высокоорганизованными животными - птицами и млекопитающими. Благодаря теплокровности и высокоразвитому мозгу они оказались лучше приспособленными к новым условиям и вытеснили менее развитых рептилий. В то же время следует отметить, что проблема массового вымирания мезозойских рептилий еще далеко не решена.