Транспорт веществ через биологические мембраны, классификация.
Первый вариант классификации - прямой и опосредованный транспорт.
Прямой - без участия переносчиков, опосредованный - с их участием. Например, перенос глюкозы с участием переносчика. Опосредованный транспорт осуществляется с затратой энергии (активный транспорт) или без затраты энергии (облегченная диффузия). Прямой транспорт всегда идет по типу пассивного транспорта.
Второй вариант классификации - это выделение двух основных видов транспорта веществ - пассивного и активного.
Третий вариант классификации - транспорт с изменением архитектуры мембраны (экзоцитоз, эндоцитоз) или без изменения структуры мембраны (все остальные виды транспорта).
Четвертый вариант классификации - это транспорт, сопряженный с переносом двух веществ (котранспорт), который может протекать по типу симпорта (два вещества идут в одном направлении - например Na + глюкоза) или по типу антипорта (одно вещество идет в клетку, второе - из клетки или наоборот - Na и К).
Антипод котранспорта - обычный транспорт, или унипорт, т. е. когда переносится одно вещество, например, молекулы глюкозы.
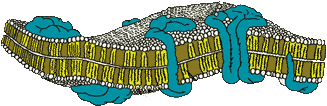
Мембрана нейрона, как и наружная мембрана любой клетки, имеет в толщину около 5 нм и состоит из двух слоев липидных молекул (греч. lipos жир - жироподобные вещества нерастворимы в воде), упорядоченных таким образом, что их гидрофильные концы (связывающие воду) обращены в сторону водной фазы, находящейся внутри и снаружи клетки, а гидрофобные концы (не способные растворяться в воде или смачиваться водой) повернуты в сторону от водной фазы и образуют внутреннюю часть мембраны. Липидная часть мембраны приблизительно одинакова у клеток всех типов. Что делает одну мембрану отличной от другой, так это специфические белки, которые связаны с мембраной тем или иным способом. Белки, которые фактически встроены в двойной липидный слой, называются внутренними белками. Другие белки, периферические мембранные белки прикреплены к мембранной поверхности, но не являются неотъемлемой частью ее структуры. В связи с тем, что мембранные липиды — жидкости, даже внутренние белки часто могут свободно перемещаться с места на место путем диффузии. Однако в некоторых случаях белки жестко закрепляются с помощью вспомогательных структур.
Мембранные белки всех клеток распадаются на пять классов: насосы, каналы, рецепторы, ферменты и структурные белки. Насосы расходуют метаболическую энергию для перемещения ионов и молекул против концентрационных градиентов и поддерживают необходимые концентрации этих молекул в клетке. Заряженные молекулы не могут пройти через сам двойной липидный слой. Избирательные пути для диффузии специфических ионов обеспечивают белковые каналы. Клеточные мембраны должны узнавать и прикреплять многие типы молекул. Эти функции выполняют рецепторные белки, которые представляют собой центры связывания, обладающие высокой специфичностью и сродством.
Ферменты размещаются внутри мембраны или на ней, чем облегчается протекание химических реакций у мембранной поверхности. Наконец, структурные белки обеспечивают соединение клеток в органы и поддержание субклеточной структуры. Эти пять классов мембранных белков не обязательно взаимно исключают друг друга. Так, например, тот или иной белок может быть одновременно и рецептором, и ферментом, и насосом.
Мембранные белки — это ключ к пониманию функций нейрона, а, следовательно, и функций мозга. Поскольку они занимают такое центральное место в современных представлениях о нейроне, есть смысл привести описание ионного насоса, различных типов каналов и ряда других белков, которые в совокупности наделяют нейроны их уникальными свойствами. Общая идея состоит в том, чтобы суммировать важные характеристики мембранных белков и показать, как эти характеристики определяют нервный импульс и другие сложные особенности функций нейрона.
Подобно всем другим клеткам нейрон способен поддерживать постоянство своей внутренней среды, заметно отличающейся по составу от окружающей его жидкости. Особенно поразительны различия в концентрациях ионов натрия и калия. Наружная среда приблизительно в 10 раз богаче натрием, чем внутренняя, а внутренняя среда примерно в 10 раз богаче калием, чем наружная. Как калий, так и натрий способны проникать через поры в клеточной мембране, поэтому некоторый насос должен непрерывно производить обмен вошедших в клетку ионов натрия на ионы калия из наружной среды. Такое выкачивание натрия осуществляется внутренним мембранным белком, называемым Na-K-аденозинтрифосфатазным насосом, или, как его чаще называют, натриевым насосом.
Белковая молекула натриевого насоса (или комплекс белковых субъединиц) имеет молекулярный вес около 275000 и размеры порядка 6x8 нанометров, что несколько больше толщины клеточной мембраны. Каждый натриевый насос может использовать энергию, запасенную в форме фосфатной связи в аденозинтрифосфате (АТФ), для того, чтобы обменять три иона натрия внутренней среды клетки на два иона калия наружной среды. Работая с максимальной скоростью, каждый насос способен транспортировать через мембрану около 200 ионов натрия и 130 ионов калия в секунду. Однако фактическая скорость регулируется в соответствии с потребностями клетки. У большинства нейронов имеется от 100 до 200 натриевых насосов на квадратный микрон мембранной поверхности, но в некоторых участках этой поверхности их плотность почти в 10 раз выше. Типичный мелкий нейрон имеет, по-видимому порядка миллиона натриевых насосов, способных перемещать около 200 миллионов ионов натрия в секунду. Именно трансмембранные градиенты натрия и калия обеспечивают возможность проведения по нейрону нервного импульса.