Лекция жоспары
1. Биологиялық физика пәні және оның басқа ғылымдармен байланысы.
2. Биологиялық мембрананың (БМ) қызметі.
3. Мембрананың құрылысы.
4. БМ липидтер мен ақуыздардың түрлері.
5. БМ фазалық ауысулар.
6. Жасанды БМ.
§ 1.1 Биологиялық физика пәні туралы.
Бізді қоршаған материалдық дүниедегі барлық құбылыстар мен өзгерістер өзара тығыз байланыста екендігі белгілі. Соның бірі айғағы ретінде биология ғылымының биологиялық жүйелердегі құбылыстарды танып білуде дәл ғылымдардың заңдылықтары мен ондағы түсініктерді қолдану арқылы ғана одан ары дамитындығына көз жеткізді. Осындай себептерден биология және физика ғылымдарының түйісінде биофизика атты жаңа ғылыми бағыт пайда болды.
Биологиялық физика биологиялық жүйені субмолекулалық, молекулалық, жасушалық, ұлпалық, мүшелік және ағзалық деңгейлердегі дамуын физика- химиялық тұрғыдан зерттейтін ғылым. Биофизика термині алғаш рет 1892 жылы Карл Пирсонның «Ғылым грамматикасы» атты ғылыми еңбегінде «...биологиялық фактілер- морфология, эмбриология және физиология физикалық заңдылықтардың дербес бір көрінісі, сондықтан оның этиологиясы...» деп жазды. Кейін А.Фик және немістің басқа да ғалымдары бұл саланы «Медициналық физика» деп атаған.
1961 жылғы І-Дүниежүзілік биофизиктердің халықаралық конгресінде биофизика өз алдына жеке ғылым ретінде танылды. Осы конгрестің шешіміне сәйкес биофизика – молекулалық биофизика, жасуша биофизикасы, күрделі жүйелер биофизикасыдеген бөлімдерден тұрады.
1. Молекулалық биофизика бөлімі- биологиялық макромолекулалар (ақуыздарды, нуклейн қышқылдарын т.б.) мен молекулалық деңгейдегі биологиялық жүйелердің құрлысы мен физикалық қасиеттерін зерттейді. Биофизиканың бұл бөлімінің пайда болуына Л.Полингтың ақуыздың кеңістіктік құрылымын мен Ф.Криктің «өмір спиралы»ДНК анықтауы үлкен себеп болды. Сонымен қатар молекулалық биофизика молекулалық деңгейде энергияның түрленуі мен миграциясын да қарастырады.
2. Жасуша биофизикасы- жасауша мен оның органелдерінің құрылымын, ондағы физика-химиялық үрдістерді, биопотенциалдың пайда болу механизмін, жасуша арқылы заттарды тасымалдауды, жасуша термодинамикасын зерттейді. Биофизиканың бұл бөлімінің дамуына Бернштейннің, А.Ходжкинның, Хакслидың және т.б. ғалымдардың қосқан үлестері орасан зор.
3. Күрделі жүйелердің биофизикасы. Биофизиканың бұл бөлімі молекулалық деңгейден бастап жасушаға дейінгі деңгейдегі биологиялық жүйедегі өз ара байланыстарды зерттеумен және оны моделдеумен айналысады.
4. Мүшелердің сезім биофизикасы.Бұл бөлім көру, есту, сезу секілді тірі биологиялық құрылымдағы құбылыстарды зерттеумен, сыртқы тітіркендіргіштердің әсерінен тірі объектінің сезу мүшелерінде, рецепторларда пайда болатын энергияның электр сигналына айналу механизмін қарстырады.
Биофизика бұл аталған бөлімдерімен қатар адам ағзасынатүрлі физиалық факторлардың –шудың, механикалық тербелістің, электр тогы мен электромагниттік өрістің, иондаушы сәулелердің және т.б. әсерлерін де зерттеумен айналысады.
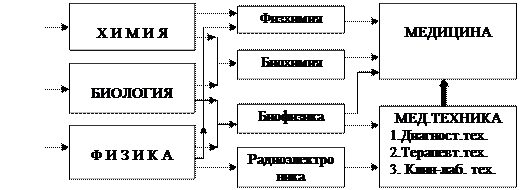
Қазіпгі кезеңде биофизика жеке ғылым ретінде физиология, биология, химия, физика, математика, компьютерлік технология, нанотехнология және т.б. ғылыми бағыттардағы ғылыми жаңалықтарды қолдана отырып және солармен бірге даму үстінде, бұл өз аралық байланыс 1- суреттегі сызбада (схемада) бейнеленген.
  |
1- сурет
Әрине, бұл схемада қазіргі замаңғы ғылыми бағыттардың барлығы ескерілді деп айтуға болмас, бірақ биофизиканың негізгі жаратылыстану ғылымдарымен өз ара байланысы көрсетілді.
Медициналық биофизика медицина және фармация ғылымдарымен бірлесе отырып мынадай мәселелерді зерттеумен айналысады:
-Азға күйін диагностикалауда жаңа физикалық әдістерді қарастыру және негіздеу (электрография, хемилюминеенция, ЭПР, ЯМР, спектроскопия т.б.).
- Тірі ағзаға сыртқы ортадағы физикалық факторларының зиянды әсер ету механизімін зерттеу (механикалық діріл, салмақсыздық, радиосәулелер т.б.)
Биофизикалық зерттеулерде физикалық, физика-хиимиялық әдістер кеңінен қолданылады. Оларға:
-рентген сәулесінің дифракциялық кескіні арқылы зат құрылысын анықтауға арналған рентгендік құрылымдық талдау әдісі;
- электрон ағынын пайдаланып зат құрылысын көруді қамтамасыз ететін электрондық микроскопты қолдану;
- зертелінетін заттардың түрлі сәулелерді жұты мен шығаруына негізделген спектроскопия әдісі.
- зертелінетін заттардың радиодиапазондағы электромагниттік толқындарды таңдамалы түрде жұтуына негізделген ЭМР және ЯПР радиоспектроскопиялық әдістер
Қазіргі заманда іргелес ғылымдардың бір- бірін қолдап, сол арқылы одан ары дамуы кең түрде байқалуда, бұл ретте биофизиканы физиканың, химияның және биологияның яғни іргелес үш ғылымның жиынтығы деп қарастырған дұрыс.
§ 1.2 Биологиялық мембрана құрылысы және қызметі.
Биофизиканың міндеттерінің бірі ретінде биологиялық мембрананың құрысысын, ол арқылы зат тасымалдауды, нерв импульсының пайда болу мен оның таралуын, рецепция құбылысын және биомембрандағы энергия өзгеруін зерттеуді атауға болады.
Мембрана биофизикасы – медицина үшін ең маңызды жасуша биофизикасының негізгі бөлімі болып саналады. Адам ағзасындағы көптеген үрдістер биологиялық мембрана арқылы жүреді және іске асады. Мембранадағы үрдістердің бұзылуы-көптеген патологиялық өзгерістердің негізгі себебі болып табылады. Адам ағзасына тигізетін емдік әсерлер биологиялық мембранаға әсер ету арқылы іске асырылады. Тіршіліктің ең кіші бөлігі болып саналатын, өз бетінше өмір сүре алатын жасуша кез келеген жәндік пен өсімдік дүниесінің негігі бөлігі болып табылады.
Биологиялық мембрана мынандай қызметтер атқарады:
- Механикалық, бұл арқылы жасуша өзін қоршаған ортадан оқшауланады, оның дербес болуына және тиісті қызмет етуіне мүмкіндік береді;
- Тосқауылдық, бұл арқылы жасуша өзін қоршаған ортамен талғампаздық (селективті) түрде пассивті және активті зат тасымалдауға қол жеткізеді;
- Матрицалық, бұл арқылы биологиялық мембранада ақуыздар мен ферменттердің болуы қамтамасыз етіледі,
- Энергетикалық, бұл арқылы биолгиялық мембранада АТФ синтезделуі, биопотенциалдардың пайда болуына мүмкіндік алады.,
- Рецепторлық, мұндай қызмет арқылы мембрана басқа жасушаларды, сыртқы тітіркендіргіштердің әсерін сезуге, заттарды танып білуге мүмкіндік алады.
Мембрана жасушыларды бір- бірінен бөлумен қатар, оның ішкі бөлігі цитоплазманы өз беттерінше жеке қызмет атқара алатын бөліктерге-компартменттерге бөледі. Қазіргі кезеңде биологиялық жүйлердің қызметін сипаттауда осы компартментализация принципі үлкен қолдау табуда. Жасушаның компартментлизациялануы, ондағы биохимиялық процесстер мен түрлі реакциялардың бір бірінен оқшау жүруіне мүмкіндік береді, бұл реакциялардың бір ортада жүруі мүмкін емес. Осы принципке сәйкес, цитоплазмада - май қышқылдары синтезделсе, олардың тотығуы-митохондрияда жүреді, ал рибосомда- ақуыз синтезделсе, оның бұзылуы-лизосомда орын алады.
«Мембрана» термині ХІХ ғасырдың орта бөлігінде пайда болды және бұл арқылы жасушаны қоршаған ортадан бөліп тұрған, жартылай өткізгішік қасиеті бар жұқа қабатты атаған. 1851 жылы физиолог Х.Моль өсімдік жасушасы плазмолизін зерттеп, оның қабырғасының мембранаға тән қасиеті бар екендігін анықтаған. 1855 жылы ботаник К.Негели жасушаның толық қанды өмір сүруіне мембрананың жартылай өткізгіштік қасиетінің маңызды екендігін, осы арқылы жасуша ішінде осмостық қысымың қалыпты жағдайда болатындығын мәлімдеді. 1890 жылы неміс зерттеушісі В.Пфеффер алғаш рет «жасуша немесе плазмалық мембрана» терминін енгізді.
Жасушаның басты құрылымы мембрана екендігін алғаш ботаниктер анықтағандығы белгілі, өйткені ХІХ ғасырда электрондық микроскоптың болмауы себепті тірі организмдегі мембрана қабатын көру әлі мүмкін емес еді. Осы себепті биологиялық мембрана құрылысы мен оның көптеген қасиеттерін анықтау ХХ ғасырдың еншісіне тиді.
Биологиялық мембрананың алғашқы үлгісі 1902 жылы ұсынылған болатын. Зерттеушілер фосфолипидте жақсы еритін заттардың мембрана қабаты арқылы еркін өтетіндігін байқаған, осы құбылыс мембрана фосфолипидтен тұратын жұқа қабат деген жорамал жасауға мүмкіндік берген.
1925 жылы Гортер мен Грендель гемолизденген эритроциттен ацетон арқылы липидті бөліп алған. Алынған ертіндіні судың бетіне құйған, булану нәтижесінде су бетінде пайда болған липид молекулаларының алып жатқан аймағының ауданы, тәжірибе басында алынған эритроциттердің ауданынан екі есе көп болған. Осының негізінде мембранадағы липидтер екі қабат болып орналасқан деген қорытынды жасалды (2- сурет).
1935 жылы бұл ойдың дұрыстығын Коул мен Кертис мембрананың электрлік параметрлерін өлшеу арқылы дәлелді. Олар мембрананың меншікті электрлік сиымдылығының С= 0,5×10-2 Ф/м2, ал меншікті электрлік кедергісінің r =10 Ом× м2 үлкен болатындығын анықтады.
Биологиялық мембрана жазық конденсаторға ұқсас. Конденсатордың екі астары ретінде мембрананың сырты және ішкі(циоплазма) орталарындағы электролиттік сұйықтарды, ал конденсатордың астарларын бөліп тұрған диэлектригі ретінде екі қатар орналасқан фосфолипид қабатын, яғни биомембараны алуға болады( 3- сурет).
 |
2- сурет 3- сурет
Жазық конденсатордың электрлік сиымдылығы мына өрнекпен анықталынады: С= e×e0×S/d, мұнан конденсатордың меншікті сиымдылығы (бірлік ауданға сәйкес келетін сиымдылық мәні) мынаған тең болады: СМС = e×e0/d, мұндағы e- липидттің диэлектрлік өтімділігі, ол e=2 тең, e0 = 8,85×10-12 Ф/м, d- конденсатор астарының ара қашықтығы. Мембрананың конденсаторға ұқсастығынан d- шаманы мембрана қабатының қалыңдығына тең деп санауға болады. Олай болса меншікті сиымдылық өрнегінен: d = e×e0/СМС. Сонда d = 8,85×10-12 Ф/м×2/0,5×10-2 Ф/м2 = 35×10-10 м = 3,5 нм., анықталған бұл шама бір қатар липид қабатының қалыңдығы.
1935 жылы Дж.Даниэлли мен Г.Давсон биологиялық мембрананың «бутерброд» тәрізді мембрананың жобасын ұсынды. Ол үш қабаттан, яғни ортасында екі қатар(биқатар) болып липидтер орналасқан, бұл қабаттың екі жағына ақуыздар жабысып орналасқан (4-сурет). Кейін бұл жобаны одан ары дамытқан көптеген ұсыныстар болды 1964 ж. Дж.Робертсон (5 сурет); 1966 ж. Дж.Ленард және С.Сингер т.б.
 |
4 сурет 5 сурет
Қазіргі уақытта 1972 жылы Никольсон мен Сингер ұсынған, 1981 жылы Сингер одан ары жетілдірген биомембрананың «сұйық-мозайкалы» моделі қолданылуда. Бұл моделге сәйкес биологиялық мембрананың негізі - липидтер, олар бір біріне перпендикуля түрде, екі қатар болып орналасқан, ал ақуыздардың кейбірі липид қабатына жабысып, кейбірі оған батып немесе оны толығымен тесіп орналасқан (6 сурет).
Жалпы липидтер үш түрде: фософолипид, гликолипид және стероид түрінде кездеседі. Бұлардың ішінде фосфолипидтердің үлесі басым, яғни көптеген денелердің мембрана қабаты фосфолипид молекуласынан құралған деп санауға болады.
 |
6 сурет. 1- фосфолипид биқабаты, 2- фосфолипид молекуласының гидрофильді полярлы басы, 3- фосфолипид молекуласының гидрофобты құйрығы, 4- интегралды ақуыздар, 5- гликолипид, 6- перифериялық ақуыздар, 7- иондық канал, 8- пор(саңлау), 9- ақуызға жабысқан микротүткіше.
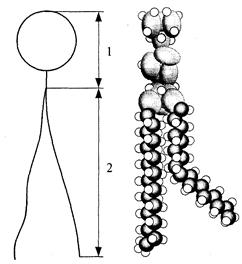
Биологиялық мембранадағы фосфолипид молекулалары екі бөліктен: полярлы бастан және полярсыз құйрықтан тұрады. Полярлы бастың алып жатқан ауданы 0,6 нм2 тең, ал құйрыққа сәйкес келетін аудан 0,2-0,3 нм2. Фосфолипид молекуласының барлық ұзындығының 1 бөлігі полярлы басқа, қалған 2 бөлігі оның құйрығына сәйкес келеді (7а - сурет).
|
|
|
|
|
а б
7 сурет
Кей фосфолипид молекуласының полярлы басы азотты топтардан (этаноламин,фосфат,холин) құралған болса, кей бірі азотсыз негіздерден (серин,инозин,треонин) құралған және зарядталған болып келеді, соның нәтижесінде ол гидрофильді қасиетке ие, осы себепті олар су молекулаларын жақсы тартады. Полярлы бас «мойын» арқылы «құйрықпен» жалғасқан. «Мойын» деп көп атомды спирттер: глицерин немесе сфингозиннен құралған құрылымды атаймыз. Құрамындағы спирт түріне сәйкес фосфолипидтерді: глицерофосфатты және сфингофосфатты деген түрлерге бөледі.
Глицерофосфатты немесе сфингофосфатты «мойынға» полярсыз құйрық жалғанған, ол екі тармақ түрінде, көміртегінің 14 - 24 атомынан тұратын, май қышқылы молекулаларынан құралған тізбек түрінде болады.
Тізбектегі көміртегі атомына бір немесе екі сутегі атомы жабысып орналасқан, құрамы мен құрылымы парафинге ұқсас, су жұқпайды, яғни гидрофобты болып келеді (7б- сурет). Екі тармақты фосфолипид құйрықшасының бірі қаныққан, екіншісі қанықпаған май қышқылынан құралады.
Фосфолипдтің физика-химиялық қасиетінің ең маңыздысы ретінде, оның басының гидрофильді, құйрықшасының гидрофобты болып келуін атаған жөн. Осындай қасиетке сәйкес және кей шарттарды сақтаған жағдайда, сулы ортада фосфолипид молекулалары өз беттерінше сфера түрінде жиналады. Нәтижесінде, фосфолипид молекулалары бір біріне паралелль екі қатар(биқатар) болып орналасады және әр қатардағы (моноқатар) молекула полярлы басымен сыртқы ортаға қарай, қышқылды құйрықшалары ішкі ортаға қарай бағытталып орналасады.
Осылайша, динамикалық жағынан тұрақты болатын фосфолипидті биқабат құрылады және ол биологиялық мембрананың негізгі қаңқасы болып саналады. Мысалы, массасы 1 мг липид молекуласынан ауданы 1 м2 болатын биқабатты жасауға болады. Жасуша мембранасының 1 мкм2 ауданында 4×106 липид молекуласы бар, бұл әр моноқабатта 2×106 молекула бар деген сөз.
Әр түрлі фосфолипид молекулаларының сулы ортада екі қатар болып жинақталу қабілеті әр түрлі, әсіресе ол лецитинде жоғары, егер ортада лецитин бар болса, ол фосфолипидтердің жиналғыштық қабілетін күшейтеді. Ғалымдар, фосфолипид молекуласының осындай қабілеті арқасында жер бетінде тіршілік пайда болған деп санайды.
Жүрізілген ғылыми зерттеулер мембрана қабатындағы липид молекулаларының тығыз орналасауы фосфолипид құрамына қандай май қышқылы енетіндігіне байланысты екендігін көрсетті. Егер көміртегі тізбесіндегі атомдар арасындағы қос байланыс көп болса, мембранадағы молекулалар арасы алшақ болады, бұл өз кезегінде мембрананың беріктілігін (қаттылығын) кемітеді және мембрана қабатының заттарды өткізгіштігін жоғарылатады.
Мембранадағы фосфолипид молекулаларының тығыз орналасуына холестериннің әсері бар екендігі анықталды. Холестерин фосфолипид биқабатына еніп, оның тығыздығын артырады. Биологиялық мембранада холестерин молекуласының көбейуі фосфолипид молекуласының алатын ауданын кемуіне алып келеді, өйткені фосфолипидтер құйрықшаларындағы көмірсутегі тізбегінің биқабатқа қатысты орналасу көлбеулігі өзгереді. Холестерин көбейген сайын фосфолипид құйрықшалары мембрана қабатына тіктеу орналасады, нәтижесінде мембрана тығыздығы одан ары артып, заттарды өткізуін кемітеді. Эритроцит плазмолемасындағы 1 моль фосфолипидке 0,37 моль холестерин келсе, мембрана ішіндегі митохондриасында бұл шама 0,03 мольге тең. Жасушадағы холстериннің щамасы ағзадағы зат алмасуға тікелей байланысты, ол өз кезегінде тамақтану түріне тәуелді. Сондықтан, холестерині көп тамақты үнемі пайдалану жасушаның қаттылығын арттырып, оның заттарды өткізу қабілетін төмендетеді.
Биологиялық мембрананы құрайтын гликолипидмолекуласы қатар жатқан жасушалардың бір біріне жабысып қалмауын қамтамасыз етеді. Бұл липидтер қатар жатқан жасуша беттерінде теріс электр зарядтарын туғызады, нәтижесінде пайда болған электростатикалық өріс әсерінен олар бір бірінен алшақтайды. Егер мембранадағы гликолипидтер белгілі бір шамадан асып кетсе, онда жасушалар бір біріннен тіптен алшақтап кетеді, нәтижесінде олардың өз ара ақпарат алмасуы бұзылады.
Зерттеулер биологиялық мембраналардың липидтік құрамы түрлі болатындығын анықтады (1 кесте).
1 кесте.
Биологиялық мембрананың липид құрамы,%
| Мембрана липидтері | Жануар жасушасының мембранасы | Е.coli бактериясының мембранасы | ||
| миелинді қабықша | эритроцит тің плазмо леммасы | ішкі митохон дриа мембрансы | ||
| Холестерин | ||||
| Фосфатидилэтанол амин | ||||
| Фосфатидилсерин | ||||
| Фосфатидилхолин | ||||
| Фосфатидилин озитил | ||||
| Сфингомиелин | ||||
| Цереброзид |
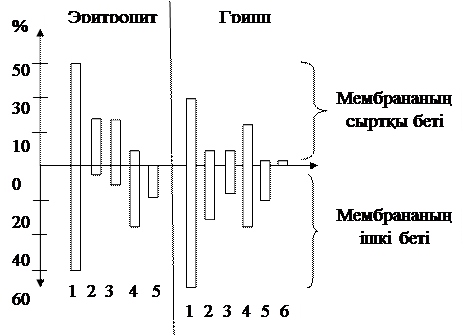
Сонымен қатар мембрана биқабатының ішкі және сыртқы беттеріндегі липид мөлшері де түрлі деңгейде болады, оны мына 8 суреттен байқауға болады.
 |
8 сурет.
1-фосфолирид, 2-сфигомиелин, 3-фосфатидилхолин,
4- фосфатидилэтаноламин, 5- фосфатидилсерин, 6- фосфатидилинозитил.
Сыртық фактолардың әсерінен липид қабаты үздіксіз өзгерістерге ұшырап отырады. Бұл құбылыстьың бір көрінісі ретінде фосфолипид құйрықшасының май қышқылды бір тармағының еркін радикалды тотығу әсерінен бұзылатындығын атуға болады, сонымен қатар фосфолипаза А2 ферменті әрекетінен қанықпаған май қышқылды тармақ жұлынып кететіндігін де атуға болады. Егер ағзаға енетін фосфолипаза А2 қажетті мөлшерден артып кетсе, мысалы улы жылан шаққан кезде, онда мембрана жасушасының бұзылуы өте жоғары деңгейге жетеді, бұл өз кезегінде ағзаның өмір сүруін тоқтауға дейін алып келеді. Микро ағзалардан бөлінетін кей ферменттер мысалы, фосфолипаза С, мембрананың липид қабатындағы фосфолипид молекуласының полярлы басын «жұлып» алады екен.
Сондықтан фосфолипаза А3 ферменті, фосфолипаза С, еркін радикалды тотығу үрдісінің мембрананы зақымдайтын әрекеті бар екендігін білу қажет.
§ 1.3 Биологиялық мембрананың физикалық қасиеттері
Тығыздығы.Фосфолипид биқабатының тығыздығ 800 кг/м3 тең, бұл судың тығыздығынан кем.
Өлшемі. Электрондық микроскопиялық талдау нәтижесінде мембрана қалыңдығы 4 нм ден 13 нм дейінгі аралықта болатындығы және жасуша мембраналардың қалыңдығы олардың түрлеріне байланысты болып келетіндігі анықталды.
Мықтылығы. Мембрананың сыртқы әсерлерге (үзілу, созылу,бұзылу) төзімдігілі төмен. Мысалы, мембрана ішіндегі қысым 100 Па болғанда мембрана құрылымы бұзылады. Тірі жасуша осмостық қысымға төтеп беру үшін мембранасын созбайды, ол өз пішінін (формасын) өзгерту арқылы төтеп береді.
Деформациялануы.Жасуша мембранасының деформациялануы мен иілгіштігі жеңіл, ал оның серпімділік модуль 0,45 Па тең.
Тұтқырлығы. Мембараның липид қабатының тұтқырлығы 30-100 мП×с, яғни өсімдік майының тұтқырлығына жақын.
Беттік керілуі 0,03-3 мН/м аралығында жатыр, бұл сумен салыстырғанда 2-3 есе кем.
Өткізгіштігі. Биомембрананың суды өткізу көрсеткіші 25-33×10-4 см/с аралығында.
Электр сиымдылығы. 1 см2 мембрана қабатының электрлік сиымдылығы 0,5 -1,3 мкФ аралығында, ал мембрана қабатындағы электр өрісінің кернеулігі Е =20×106 В/м. Мембрананың фосфолипид аймағының диэлектрлік өтімділігі e = 2,0-2,2 тең болса, оның гидрофильді аймағының диэлектрлік өтімділігі e = 10-20 тең.
Электрлік кедергісі. 1 см2 мембрананың бетінің кедергісі 102 – 105 Ом/см2, ол жасушаның сыртқы немесе ішкі орталарымен салыстырғанда ондаған миллион есе көп. Мембананың электро оқшаулау (электроизоляциялау) қасиеті техникалық изолятордан әлде қайда көп жоғары.
§ 1.4 Мембрананың фазалық күй.
Бір қалыпты физиологиялық жағдайда, яғни температура адам денесінің температурасына тең, рН және иондар концентрациясы тұрақты болғанда биологиялық мембрана қабаты «сұйық кристаллды» күйде болады. «Сұйық кристалл» күй деп құрылымда сұйыққа да (құрылым бөлшектері ретсіз, хаосты қозғалыста), кристаллға да (құрылым бөлшектері кеңістікте реттелген күйде) тән қасиет бір мезгілде кездесетін күйді атаймыз.
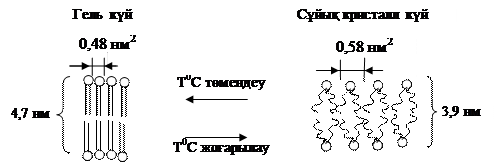
Температура төмендегенде биомембрананың фосфолипидтік қабаты өз құрылымын сақтай отырып қатты кристалл күйге көшеді. Мембрана қабатындағы фосфолипид молекулаларының тығыздығы оның күйіне тікелей тәуелді, мысалы, лецитин молекуласы қатты кристалл күйде 0,46-0,48 нм2 аймақты алса, оның сұйық кристалл күйдегі алатын аймағы 0,6-0,8 нм2 тең, мұнан мембрана қабатының өлшемдері (көлемі, ауданы, қалыңдығы т.б.) оның фазалық күйін тікелей тәуелді деген қорытынды жасауға болады. Фосфолипид молекуласының құйрығындағы қанықпаған май қышқылдарындағы қос байланыс көп болған сайын, қатты кристалл күйге ауысу температурасы да төмендейді.
Сұйық кристаллды биомембрана құрылымы температура өзгерісіне өте сезімтал. Температура төмендегенде мембрана сұйық кристалл күйден гель тәрізді күйге ауысады. Бұл кезде мембрана өзінің толық құрылымын сақтайды, фосфолипид құйрышқалары түзуленіп, бір біріне параллель орналасады және олардың тербелісі шектеледі. Сұйық кристалл күйде бір липид молекуласы алып жатқан аудан 0,58 нм2 болса, гель күйде бұл шама 0,48 нм2 дейін төмендейді, яғни мембрана көлемі азаяды, оның есесіне мембрана қабаты қалыңдайды(9- сурет)
 |
9- сурет
Биомембрана кабатының сұйық кристалл күйден гель күйге көшуі кезінде биқабатта радиусы 1-3 нм болатын порлар(саңлау) пайда болады. Бұл порлар арқылы иондар мен төменгі молекулалы қосылыстағы заттар тасымалданады, яғни фазалық өзгеріс мембранының иондық өткізгіштігін жоғарылатады. Мұндай қасиеттің арқасында, яғни дене температурасының төмендеуі кезінде мембраналардың зақымдалуы болмайды, өйткені су және тұздардың мембрана арқылы өтуі жоғарылап, олардың жасуша ішінде кристаллдануына жол берілмейді, бірақ мұндай күйде липид молекулаларының қозғалғыштығы төмендейді.
Мембрана қабатында ақуыздар көп, олардың көптігінің әсерінен мембрананың беттік керілу көрсеткіші «липид-су» шекарасындағы көрсеткіштен «ақуыз-су» шекарасындағы көрсеткішіне жақын. Мембаралық ақуыздардың концентрациясы жасуша түріне байланысты өзгеріп отырады, мысалы, аксонның миелинді қабатындағы ақуыз липидтерден 2,5 есе аз, ал эритроцит мембранасындағы ақуыз саны керісінше липидтен 2,5 көп.
Ақуыздар мембрана қабытында әр түрлі орналасқан, бірі мембрана бетіне жабысып тұрса, бірі оны тесіп өтеді. Мембрана бетіне жабысып орналасқан ақуыздарды перифериялық, ал оны тесіп өткендерін интегралдық ақуыздар беп атайды.
Интегралды ақуыздар липиттер тәрізді, мембрана қабытындағы бөлігі a-спираль түрінде ширатылған түрде болып келетін гиброфобты қасиеті бар, аминоқышқылдарынан тұрады, ал мембрана қабатынан сыртқа шыққан бөлігі гидрофильді қасиетке ие және аминоқышқылдарынан тұрады. Бұл ақуыздар мембрана қабытына гидрофильді әсерлесу нәтижесінде пайда болатын күштер арқылы ұсталып тұр. Кейбір интегралды ақуыздардың мембранадан сыртқа шыққан бөлігіне көмірсулар жабысып тұрады, мұндай комплекстерді гликопротеин деп атайды, олар рецепторлық қызмет атқарады және ақзаның иммундық реакцияларында маңызды роль бар. Мысал ретінде мембрана қабатын 7 рет тесіп өткен бактериородоспинді атауға болады, ол ақуызға фоторецепторлық қабілет береді, ал эритоцит ақуызы - гликофорин қан тобын анықтайтын гликопротейнге жатады. Көптеген интегралды ақуыздар мембрана қабатында канал ролін атқарады, мұндай каналдардың ішкі бетінде гидрофильді аминқышқылының радикалыдары орналасады, олар арқылы суда еритін молекулалар мен иондар өтеді.
Перифериялық ақуыздар мембрананың сыртқы бетіне де, ішкі бетіне де орналаса алады. Бұл ақуыздар мембрана қабытына электростатикалық әсерлесу нәтижесінде пайда болатын күштер арқылы ұсталып тұр, бұл күштер интегралды ақуыздарды ұстайтын күшпен салыстырғанда әлде қайда төмен, сондықтан перифериялық ақуыздарды мембрана бетінен жұлып алу жеңіл. Ал интегралдық ақуыздар мембрана қабатындағы липидтердің көмірсутегі тізбегімен Ван-дер Ваальс күші арқылы әсерлеседі және бұл әсерлесу күшінің үлкен болуы себепті интегралды ақуызды бөліп алу тек липид қабатын бұзу нәтижесінде қол жеткізуге болады.
Мембрана құрамындағы липидтер мен ақуыздар қозғалғыш келеді, егер молекула қозғалысы мембрананың бір қабатында орын алса, ондай қозғалысты – латериалды диффузия деп атайды, егер молекула мембрананың бір қабытынан екінші қабаттына орын ауыстырса, оны флип-флоп орын ауыстыру деп атайды.
Латериалды диффузия кезіндегі молекуланың бір орыннан екінші орынға ауысу жиілігі мынаған тең:
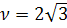
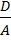 ,
,
мұндағы D – латериалды диффузия коэффициенті, A- мембрана бетіндегі бір молекула алып жақтан бет ауданы.
Молекуланың бір орындағы «тұрғылықты өмір ұзақтығы» оның орын ауыстыру жиілігіне кері пропорционал болады:
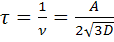 ,
,
Молекуланың t уақыт мерзіміндегі орташа квадраттық орын ауыстыруы мына өрнекпен анықталынады:
 =
=  ,
,
Липидтердің диффузия коэффиценті өте жоғары. Мысалы, 400С температура кезіндегі ретикулум саркоплазмасының липидтері үшін D =1,2×10-11 м2/c тең. Егер А= 7×10-19 м2 болса, онда липидтер 1 секунд ішінде 5,9×107 рет орын ауыстырар еді, мұнан липидтің «тұрғылықты өмір ұзақтығы» 1,7×10-8с тең болатындығы көрінеді.
Ақуыз молекулалары липдтер тәрізді жылдам қозғалмайды. Мысалы, лимфоцитарлы мембрананың антиген молекуласының ақуызы үшін D =10-14 м2/c тең болса, онда оның орташа квадраттық орын ауыстыруы 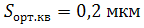 тең болады. Мұнан мембранадағы ақуыздардың қозғалшыштығы төмен екендігін көреміз, өйткені мембрана қабатындағы микротүтіктер немесе микрофиламенттер ақуыздардың басым бөлігінің қозғалысын шекетеп, оларды мембрананың белгілі бір бөлігінде тұруына ықпал етеді.
тең болады. Мұнан мембранадағы ақуыздардың қозғалшыштығы төмен екендігін көреміз, өйткені мембрана қабатындағы микротүтіктер немесе микрофиламенттер ақуыздардың басым бөлігінің қозғалысын шекетеп, оларды мембрананың белгілі бір бөлігінде тұруына ықпал етеді.
Мембрана молекулалары түзу сызықты қозғалыспен қатар айналмалы қозғалысқа да түседі. Мысалы, 1 радин бұрышқа фосфолипид молекуласы 10-9 секунд ішінде бұрылса, родоспин 10-6 с, цитохромоксидаза 10-4 с ішінде бұрылады.
Мембаранадағы фофолипид молекуларладың флип-флоп түріндегі қозғалысы латериалды диффузиямен салыстырғанда өте баяу жүреді, мысалы, фосфолипид молекуласы мембрананың бір қабатынан екінші қабатына өтуіне 1 сағатқа жақын уақыт қажет екен. Ал ақуыз молекулалары мұндай орын ауыстыруға қатыспайды.
Латериалды және флип-флоп диффузия жылдамдықтардың әр түрлі болуының үлкен маңызы бар. Латериалды диффузия жылдамдығының үлкен болуы мембранадағы химиялық рекациялардың жылдам өтуіне ықпал етеді, ал флип-флоп орын ауыстыруының баяу өтуі мембранадағы тепе теңсіздікті қамтамасыз етеді.