Скафолдні і адапторні білки
Характерною рисою молекул цього типу є відсутність ферментативної активності при одночасній наявності доменів, які відповідальні за взаємодію з іншими білками, а також залишків тирозинів, які можуть бути фосфорильованими. Роль цих молекул у нашому випадку- це посередництво між поверхневими рецепторами і білками, що ведуть до активації різних шляхів трансдукції сигналів. Скафолдні білки є більшими, мають ряд сайтів для тирозин-залежного фосфорилювання, з подальшим рекрутуванням ряду білків. Адапторні білки менші, мають 2 чи 3 домена для зв’язування інших білків (SH2, SH3). До великих скафолдних білків належать – LAT, SLP-76, BLNK та малі адаптори- GADS , Grb 2 та інші. Так, сигнальний білок SOS має збагачені проліном мотиви, які взаємодіють з SH3 доменами Grb2.
 |
II. Малі G-білки - група G-білків – родина малих ГТФ- зв’язуючих білків (напр., Ras) – є мономерами (в даному випадку мономер 21 кДа) і беруть участь у передачі сигналу від рецепторів із тирозинпротеїнкіназною активністю, що належать до Single-TMS Receptors (1-ТМS-рецепторів).
• Неонкогенні форми (с- Ras) є в усіх клітинах. Вони є регуляторами їх росту й диференціації, залучаються у стимуляцію клітинних поділів факторами росту. Ras активує протеїнкінази Raf й фосфатидилінозитол -3-кінази. Raf, в свою чергу, є першим членом каскада кіназ, який спричиняє активацію ЕRК і веде до транскрипції генів.
• Всі члени мають певну гомологічну послідовність й поділяються на підродини - Ras, Rho, Raс, Cdc42. В межах кожної підродини спостерігається більш сильна гомологія. Відомо понад 70 таких білків. Всі вони є одноланцюговими поліпептидами, що заякорені в плазматичній мембрані клітин.
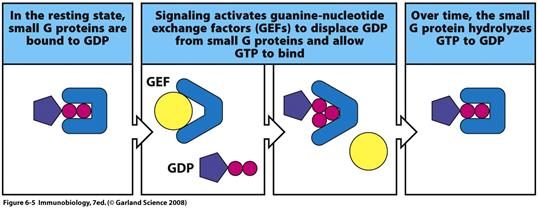
Всі члени цієї родини характеризуються подібною первинною структурою та організацією функціонального центру, що дає можливість припустити, що всі вони походять від спільного попередника. Більшість G –білків можуть утворювати стабільні комплекси зі своїм субстратом GTP і продуктом реакції - GDP. Висока спорідненість до гуанілових нуклеотидів і низька (як правило) швидкість самовільного гідролізу ГТФ необхідні для уповільнення спонтанного переходу між конформаціями зі зв’язаним ГТФ чи ГДФ. В активному ГТФ-зв'язаному стані G -білок передає сигнали далі в клітину. Однак G - білок залишається в активному стані дуже короткий проміжок часу (сек), після чого активується його власна ГТФазна активність. Фактично цей гідроліз забезпечує механізм негативного зворотного зв’язку, який забезпечує коротку тривалість перебування G білка в активованому стані.
Для переведення білків з одного стану в інший часто є необхідною присутність білків-регуляторів. Один підклас- це білки-активатори ГТФазної активності (GАP-білки= GTP-ase activating protein),які зв’язуються з G- білками на певних етапах їх функційного циклу та беруть участь у формуванні активного центру, або стабілізують конформацію G-білка в активному стані. Інший клас регуляторних білків-GEF –білки=GEF-guanine nucleotide exchange factor, які необхідні для прискорення обміну ГДФ на ГТФ в активному центрі G-білка.
 |
На прикладі системи Ras-білків можна наглядно представити схему функціонування класичного G-білка.Ras-білки- мембранно-зв'язані глобулярні білки, які виконують роль молекулярного перемикача, координуючи регуляторні сигнали між клітинною поверхнею і ядром.
Беруть участь у процесах клітинного поділу і диференціації.
Ras-білки існують в неактивній ГДФ-зв'язаній формі (off)-виключено і в ензиматично -активній ГТФ-звязаній формі (on) - включено.
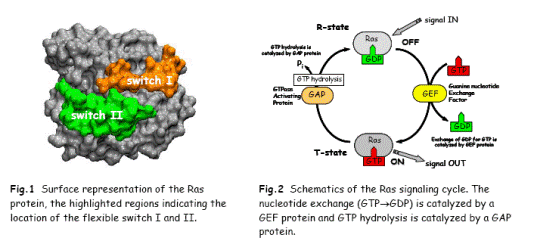
В основі структури кожного G-білкалежить ГТФ зв’язуючий домен, що складається з 6 β –складок, які оточені α-спіралями.
Ділянки, що беруть участь у зміні конформації (рухомості) при гідролізі ГТФ, представлені двома switch-сайтами ( сайтами-перемикачами) : I- ам-тні залишки 30-38 та II- ам-тні залишки 60-76. В стані спокою Ras-білки мають ГДФ-зв'язуючий статус. Після надходження сигналу Ras активується при обміні ГДФ на ГТФ. Цей процес каталізується GEF-факторами ( для Ras- ГТФаз –Sos). Трансдукція сигналу йде той час, коли Ras-білки знаходяться в активній ГТФ-зв'язаній формі. Після взаємодії з GАP-білками відбувається підвищення власноїГТФазної активності Ras-білків в 105 раз! Для цієї системи показано, що Ras GАP беруть участьв стабілізації активного центру ферменту в перехідному стані за рахунок компенсації частково негативного заряду на фосфаті позитивно зарядженим залишком аргініну і тим самим каталізувати проходження реакції гідролізу ГТФ. До Ras GАPналежать NF1- продукт гена, що бере участь у нейрофіброматозі та 120 kDa Rasподібний GАP.
Втрата можливості фактично виключити Ras-білки шляхом взаємодії з GАPпризводить до неконтрольованої проліферації, а саме раку. Вважається, що активована форма Ras генів відповідає за 30% ракових захворювань людини.
 |
III. В результаті активації рецепторів відбувається і активація фосфатидилінозитолкіназ, що фосфорилюють мембранні фосфатидилінозитоли. Так активація ФІ3-кінази, яка каталізує утворення фосфатидилінозитол-3,4,5-трифосфату (ФІФ3) з ФІФ2(фосфатидил-4,5-біфосфату), приводить до рекрутування до мембрани ряду сигнальних білків, таких як протеїнкінази AKT, ITK, до складу яких входять РН-домени = pleckstrin homology-зв'язування, зокрема з фосфоліпідами клітинної мембрани та з протеїнкназою С=ПКС (фосфоінозитиди).
 | |||
 |