Методические рекомендации по самоподготовке.
МЕХАНИЗМЫ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ.
ПРИНЦИПЫ КООРДИНАЦИОННОЙ ДЕЯТЕЛЬНОСТИ ЦНС
Продолжительность изучения темы_______________ часов
Из них на занятие ___________ часов; самостоятельная работа_________ часов.
Место проведения учебная комната
Цель:Знать виды и механизмы центрального торможения; Уметь оценить роль центрального торможения в координации рефлекторной деятельности организма.
Задачи:
1. Знать историю открытия центрального торможения (Шеррингтон, Сеченов, Гольц) и современные исследования, позволившие раскрыть его природу (Экклс, Реншоу);
2. Уметь перечислить основные виды центрального торможения, связанного и не связанного с функцией особых тормозных нейронов;
3. Уметь охарактеризовать сущность, механизм и основные разновидности постсинаптического торможения;
4. Знать медиаторные и ионные механизмы тормозных потенциалов (ТПСП), лежащие в основе постсинаптического торможения;
5. Уметь охарактеризовать сущность и механизм пресинаптического торможения;
6. Знать медиаторные и ионные ТПСП, лежащие в основе пресинаптического торможения;
7. Уметь охарактеризовать принципиальные возможности нейронной деятельности, способствующие ослаблению процесса возбуждения (торможение вслед за возбуждением, пессимум, оклюзия);
8. Уметь четко определить биологическое значение и возможности возникновения каждой разновидности центрального торможения;
9. Уметь рассмотреть взаимодействие процессов возбуждения и торможения как необходимое условие для наилучшего осуществления рефлекторных актов организма;
10. Знать, что в основе координации рефлексов лежат принципы и особенности распространения возбуждения и торможения в ЦНД;
11. Знать сущность наиболее важных принципов координационной деятельности ЦНС (реципрокность, обратная связь, окклюзия, облегчение, конечный путь, доминанта, субординация).
Методические рекомендации по самоподготовке.
Нервная система человека и животных может быть представлена как система нейронных цепочек, передающих возбуждающие и тормозные сигналы (нервная сеть). Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Такие нейронные цепи построены из стандартных элементов, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных нервных структур.
Существуют значительные количественные различия нервных сетей у разных видов позвоночных и беспозвоночных. Так, у человека нервная система включает около 1010 элементов, у примитивных беспозвоночных - около 104 нейронов, Однако в строении и функционировании всех нервных систем имеются общие черты. Практически во всех отделах центральной нервной системы обнаружены дивергенция нервных путей, конвергенция нервных путей и различные варианты тормозных связей между элементами нервных цепочек.
 Дивергенция и конвергенция путей. Дивергенция (расхождение) пути (рис 2.А) - возникает в результате контактирования одного нейрона с множеством нейронов более высоких порядков. Так, например, происходит разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее - на моторные нервные клетки. Дивергенция пути сигнала наблюдается так же у вставочных и у эффекторных нейронов.
Дивергенция и конвергенция путей. Дивергенция (расхождение) пути (рис 2.А) - возникает в результате контактирования одного нейрона с множеством нейронов более высоких порядков. Так, например, происходит разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее - на моторные нервные клетки. Дивергенция пути сигнала наблюдается так же у вставочных и у эффекторных нейронов.
Рис 2. Дивергенция (А), конвергенция (Б) и пространственная суммация (В) нервных путей в центральной нервной системе.
Дивергенция пути обеспечивает расширение сферы действия сигнала, благодаря ей, информация поступает одновременно к разным участкам ЦНС. Это называют иррадиацией возбуждения (или торможения). Дивергенция настолько обычное явление, что можно говорить о принципе дивергенции в нейронных цепях.
Конвергенция - это схождение многих нервных путей к одним и тем же нейронам (рис 2.Б). Например, у позвоночных на каждом мотонейроне спинного мозга и ствола головного мозга образуют нервные окончания тысячи сенсорных, а также возбуждающих и тормозных вставочных нейронов разных уровней. Мощная конвергенция обнаруживается и на нейронах ретикулярной формации ствола мозга, на многих корковых нейронах у позвоночных и, видимо, на командных нейронах.
Конвергенция многих нервных путей к одному нейрону делает этот нейрон интегратором соответствующих сигналов. Вероятность возбуждения такого нейрона-интегратора зависит не от каждого пришедшего стимула в отдельности, а от суммы и направления стимулов, действующих одновременно, то есть суммы всех синаптических процессов происходящих на его плазматической мембране. Другими словами, вероятность распространения возбуждения через нейрон-интегратор определяется алгебраическим сложением величин возбуждающих и тормозных входов на нем, активных в данный момент. Такое сложение является результатом или пространственной или временной суммации. Пространственная суммация – результат сложения нервных импульсов приходящих одновременно к нейрону через разные синапсы (рис 2.В), временная суммация – сложение приходящих поочередно, через один синапс с небольшими интервалами времени. В обоих случаях нейрон интегратор, называют общим путем для конвергирующих на него нервных сигналов, а если речь идет о мотонейроне, т. е. конечном звене нервного пути к мускулатуре, говорят об общем конечном пути.
Результат суммации заключается в возможности изменения направления распространения возбуждения в ЦНС, (то есть не строго в пределах одной рефлекторной дуги), а значит и в изменении характера ответной реакции организма в ответ на действие раздражителя. Ответ организма, реализуемый в результате, становится более адекватным внешним условиям и состоянию нервной системы. Пример такого выбора ответа можно видеть, если речь идет о конвергенции не на одном нейроне, а на группе нейронов совместно регулирующих общую функцию, что в ЦНС не редкость. Наличие конвергенции множества путей на одной группе мотонейронов лежит в основе феноменов пространственного облегчения и окклюзии.
Пространственное облегчение - это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Т.е. при раздельном действии афферентных сигналов возбуждение возникает в меньшем числе эфферентных нейронов и эффект оказывается слабее. Феномен объясняется суммацией совместно возникающих ВПСП до критического уровня деполяризации в группе мотонейронов, в которых при раздельной активации входов ВПСП оказывались слишком слабыми для генерации ответа.
Окклюзия - это явление, противоположное пространственному облегчению. В этом случае эффект окажется выше если афферентные сигналы действуют порознь, а при их совместном действии возбуждается меньшая группа мотонейронов. Причина окклюзии состоит в том, что здесь афферентные входы в силу конвергенции частично связаны с одними и теми же мотонейронами, и каждый может возбуждать их, как и оба входа вместе.
return false">ссылка скрытаТаким образом, если эффект нескольких стимулов, поступающих одновременно или в быстрой последовательности будет выше, чем сумма эффектов отдельных стимулов то это явление называется облегчением; если же эффект на сочетание стимулов меньше, чем сумма ответов на отдельные стимулы, то такое явление - окклюзия.
Такое явление следует учитывать, например, при тренировке различных функцинальных показателей скелетных мышц.
Простые тормозные и усиливающие цепи.
Тормозные цепи, виды торможения. Торможение, как и возбуждение, - активный процесс, оно возникает в результате сложных физико-химических изменений в тканях. Благодаря процессу торможения достигается ограничение распространения возбуждения в ЦНС и обеспечивается координация рефлекторных актов, внешне этот процесс проявляется ослаблением функции какого-либо органа.
Открытие торможения в ЦНС было сделано основоположником русской физиологии И. М. Сеченовым. В 1862 г. Им были проведены классические опыты, получившие название «центральное торможение». И. М. Сеченов на зрительные бугры лягушки, отделенные от больших полушарий головного мозга, помещал кристаллик хлорида натрия (поваренная соль) и наблюдал при этом увеличение времени спинномозговых рефлексов. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась. Результаты этого опыта позволили И. М. Сеченову сделать заключение о том, что в центральной нервной системе наряду с процессом возбуждения развивается и процесс торможения, способный угнетать рефлекторные акты организма.
К настоящему времени анализ тормозных явлений в ЦНС позволил выделить две формы разновидности постсинаптическое и пресинаптическое торможение.
Постсинаптическое торможение развивается не постсинаптических мембранах межнейронных синапсов и связано с гиперполяризацией постсинаптической мембраны под влиянием медиаторов, которые выделяются при возбуждении специальных тормозных нейронов. При этом, локально возникающая на постсинаптической мембране гиперполяризация - тормозной постсинаптический потенциал (ТПСП) – затрудняет электротоническое распространение возбуждающих постсинаптических потенциалов (ВПСП) от других синапсов, к аксонному холмику. В результате в зоне аксонного холмика не происходит выведение мембранного потенциала на критический уровень. Потенциал действия не образуется, нейрон не возбуждается.
Постсинаптическое торможение активно используется в нейронных сетях, и в зависимости от вариантов связывания нейронов друг с другом выделяют несколько его видов: реципрокное (прямое), параллельное, возвратное, латеральное (рис.3)
Реципрокное торможение (рис 3.А) – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов. Классический пример реципрокного торможения - это торможение мотонейронов мышц-антагонистов у позвоночных. Торможение осуществляется с помощью специальных тормозных вставочных нейронов. При активации путей, возбуждающих, например, мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами вставочных клеток.
Возвратное торможение (рис. 3.Б) - это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам. Возвратное торможение наблюдается, например, в мотонейронах спинного мозга позвоночных. Эти клетки отдают возвратные коллатерали в мозг к тормозным вставочным клеткам Реншоу, которые имеют синапсы на этих же мотонейронах. Торможение обеспечивает ограничение ритма мотонейронов, позволяющее чередовать сокращение и расслабление скелетной мышцы, что важно для нормальной работы двигательного аппарата.
Такую же роль играет возвратное торможение и в других нервных сетях.
Параллельное торможение (рис. 3.В) – играет сходную с возвратным роль, но в этом случае возбуждение блокирует само себя, посылая тормозной сигнал на нейрон который одновременно и активирует.
Это возможно, если возбуждающий импульс сам не должен вызвать возбуждения на нейроне-мишени, но его роль важна при пространственной суммации, в комбинации с другими сигналами.
Латеральное торможение (рис. 3.Г) – это торможение нервных клеток, расположенных по соседству с активной, которое этой клеткой и инициируется. При этом вокруг возбужденного нейрона возникает зона, в которой развивается очень глубокое торможение.
Латеральное торможение наблюдается, например, в конкурирующих сенсорных каналах связи. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. Во всех случаях латеральное торможение обеспечивает контраст, т. е. выделение существенных сигналов или их границ из фона.

Рис. 3. Разновидности постсинаптического торможения: А – реципрокное, Б – возвратное, В – параллельное, Г- латеральное. Темные нейроны – возбуждающие, светлые – тормозные.
Пресинаптическое торможение развивается в аксо-аксональных синапсах, образованных на пресинаптических окончаниях нейрона.
В основе пресинаптического торможения лежит развитие медленной и длительной деполяризации пресинаптического окончания, что и приводит к развитию торможения. В деполяризованном участке нарушается процесс распространения возбуждения и поступающие к нему импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделения достаточного количества медиатора – нейрон не возбуждается.
Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные нейроны, аксоны которых и образуют синапсы на пресинаптических окончаниях аксона-мишени.
Разновидности пресинаптического торможения изучены недостаточно, вероятно они те же, что и для постсинаптического торможения. Точно известно о наличии параллельного и латерального пресинаптического торможения (рис. 4).

Рис. 4. Разновидности пресинаптического торможения: А – параллельное, Б – латеральное. Темные нейроны – возбуждающие, светлые – тормозные.
В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рисунках, тем не менее, все варианты пре- и постсинаптического торможения можно объединить в две группы. Во-первых, когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение), во-вторых, когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможение).
Кроме того, тормозные клетки сами могут быть заторможены другими тормозными нейронами, это может облегчить распространение возбуждения.
Роль процесса торможения.
- Оба известных вида торможения со всеми их разновидностями выполняют, прежде всего, охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов, утомлению, истощению и прекращению деятельности ЦНС.
- Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пресинаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть полностью заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут приходить сотни и тысячи различных импульсов по разным путям, но число дошедших до нейрона импульсов определяется пресинаптическим торможением.
- Поскольку блокада торможения ведет к широкой иррадиации возбуждения и судорогам, следует признать, что торможение является важным фактором обеспечения координационной деятельности ЦНС.
Усиливающие цепи и механизмы усиления. Нейронные сети имеют не только тормозные механизмы, препятствующие распространению возбуждения, но и системы, усиливающие приходящий к ним сигнал. Рассмотрим некоторые из них.
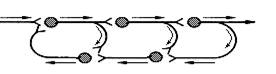 Самовозбуждающиеся нервные цепи (цепи с положительной обратной связью) (рис.5). Некоторые данные свидетельствуют о том, что в мозгу животных и человека существуют замкнутые самовозбуждающиеся цепочки нейронов, в которых нейроны соединены синапсами возбуждающего действия. Возникнув в ответ на внешний сигнал, возбуждение в такой цепочке циркулирует, иначе реверберирует, до тех пор, пока или какой-либо внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Выходные пути от такой цепочки (ответвляющиеся по коллатералям аксонов нервных клеток - участников цепи) во время работы передают равномерный поток импульсов, создающий ту или иную настройку в нервных клетках-мишенях. Ее функции могут состоять в том, чтобы обеспечивать длительное поддержание индуцированной однажды активности.
Самовозбуждающиеся нервные цепи (цепи с положительной обратной связью) (рис.5). Некоторые данные свидетельствуют о том, что в мозгу животных и человека существуют замкнутые самовозбуждающиеся цепочки нейронов, в которых нейроны соединены синапсами возбуждающего действия. Возникнув в ответ на внешний сигнал, возбуждение в такой цепочке циркулирует, иначе реверберирует, до тех пор, пока или какой-либо внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Выходные пути от такой цепочки (ответвляющиеся по коллатералям аксонов нервных клеток - участников цепи) во время работы передают равномерный поток импульсов, создающий ту или иную настройку в нервных клетках-мишенях. Ее функции могут состоять в том, чтобы обеспечивать длительное поддержание индуцированной однажды активности.
Рис.5. Самовозбуждающаяся нервная цепочка
Таким образом, самовозбуждающаяся цепочка, пока она работает, как бы «помнит» тот краткий сигнал, который включил в ней циркуляцию (реверберацию) импульсов. Считают, что это возможный механизм (или один из механизмов) краткосрочной памяти, однако этому практически нет экспериментальных доказательств.
Синаптическая потенциация — увеличение амплитуды постсинаптического потенциала, если интервал между последовательным возникновением потенциалов действия в пресинаптической мембране невелик, то есть происходит частая и ритмическая активация синапса. Явление потенциации связывают с накоплением ионов кальция в пресинаптическом окончании, который дополнительно вбрасывается туда при каждом новом стимуле и не успевает полностью удаляться между частыми стимулами. Вследствие этого, каждый новый пресинаптический потенциал вызывает высвобождение большего числа квантов медиатора.
Такую же природу имеет и посттетаническая потенциация. В этом случае увеличение числа квантов медиатора, высвобождаемых нервным импульсом, после предшествующего ритмического раздражения приводит к увеличению синаптической реакции нейрона на одиночное раздражение пресинаптических путей. Посттетаническая потенциация может длиться от нескольких минут до нескольких часов в различных структурах мозга. Предполагают, что постсинаптическая потенциация играет важную роль в пластических перестройках функций синапсов, и лежит в основе механизмов организации условных рефлексов и памяти. Например, особенно длительная посттетаническая потенциация обнаружена в гиппокампе – структуре, которая, играет важную роль в явлениях памяти и научения.
Ритмическая стимуляция может приводить и к снижению активности синапсов. Процесс снижения постсинаптических потенциалов во время или по окончании тетанической стимуляции по сравнению с исходной амплитудой называется синаптической депрессией; по аналогии с потенциацией, различают тетаническую и посттетаническую депрессию. Возможно, синаптическая депрессия имеет место во многих участках нервной системы и является нейронным коррелятом привыкания (габитуации). У беспозвоночных габитуация простых поведенческих реакций прямо соответствует депрессии участвующих синапсов; то же самое относится и к флексорному рефлексу у кошки. Таким образом, синаптическая депрессия, так же как синаптическая потенциация, составляет элементарный процесс научения.