Эписомдар.
Бактериялық клетканың хромосомына ене алатын плазмидаларды эписомалар деп атайды.Бұлардың тобына бактериофагы жатады.Оларда репликация дербес жүреді.Бактерия клеткасын талқандайдыинемесе профаг түрінде Е. Coli геномының құрамында кіреді де бірге репликация (еселенеді) жасайды. Эписома геномы өзін жеткізетін бактериялардың қасиеттерін анықтайды. Бұл қасиеттер әр түрлі болуы мүмкін, эписома көбінде бактериялардың ауру металдардың тұздарына, антибиотиктерге және басқа агенттерге төзімділігін анықтайтын ферменттерді жасап шығаруға жауапты болады. Кейде олар бактериялардың ауру туғыштығын да анықтайды. Антибиотиктерге төзімділікті анықтайтын эписомалар да кездеседі. Олар R-факторлар. Олар ішек таяқшасының бір мезгілде бірнеше антибиотиктерге төзімділігін анықтай алады және осы қасиетті бірнеше штамдарға, бактериялардың басқа түрлеріне жұқтыруы мүмкін. R-факторлар F-факторлар тәрізді жұқпалы. Олар 50-жылдары антибиотиктермен шұғылдану кезеңінде ашылды. Жапонияда дизентерия ауруларын емдеуге антибиотиктерді қолданудан кейін сипатталған ғажайып құбылысты олардың дәл осы қасиеттерімен түсіндіруге болады.Аурулар саны қасиет кеміп, содан соң қайтадан тез өсе бастады.Осы уақытта көптеген төзімділік яғни R-факторлары. Ғ-плазмидалары бар бактериялардың кездесу жиілігі де артты. Резистенттікті (төзімділікті) қамтамасыз ететін механизм R-факторлардыі антибиотиктерді бұзатын немесе инактивтендіретін фермент синтезін бақылау, клетканың өткізгіштігін антибиотик енбейтіндей етіп өзгерту қабілетімен анықталады.
ПлазмидаларПлазмидалар молекулалық массасы 1*10 Д мөлшерінде дөңгелек формасы ДНҚ малекулаларынан тұрады. Бактериялар мен эукариоттардың хромосомадан тыс генетикалық детерминанттары плазмидалардың көлемді тобын құрады. F-факторымен басқа эписомалар бактерия хромосомасында тәуелсіз өзінше екі еселенетін болғандықтан оларды да плазмидалар деп атайды.Плазмида рецепиент клеткасынан бір бөлшек ДНҚ. Түрінде клетка мембранасына бекітілген күйінде екі еселенеді. Осыдан кейін оның жабысқақ ұштары шеңбер жасап жабысады да едәуір күрделі спиральданған құрылымға айналады. Әр түрлі плазмидалардың репликациясы хромосомалық ДНК репликациясы секілді тәуелсіз реттеледі, бұл белгілі бір жағдайларда клеткада белгілі бір плазмидалардың ғана талғамды репликациялануына әкеліп соғады.
Бақылау сұрақтары:
1Рекомбинанттардың пайда болу себебі?
2Плазмидалар деген не?
3Дизентерия ауруларын не туғызады?
4Рекомбинанттардың пайда болу жиілігі?
5Плазмида малекулаларының ұзындығы?
Пайдалынылған әдебиеттер:
Инге-Вечомов С.Г.Генетика сосновами селекции М.Высшая школа 1989.
Лобашев М.Е.Ваттн К.Е.Тихомирова М.М.Генетика с основами селекция. М Просвещение 1979
Ватти К.В. Тихомирова М.М Руководства к пратическим заниятиям по генетике М.Просвещение. 1979 1972
Лобашев М.Е Генетика.Изд-во ЛГУ..Генетические 1969
Медведов И7Н.Практическая генетика М.Наука 1966
Мұхамбетжанов К.К Далабаев Б.А. Өтешева Г.А.Генетикадан практикалық сабақтар Алматы,Ғылым 2004
Мұхамбетжанов К.К Генетика.Алматы 2005
20 Дәріс
Дәрістің тақырыбы: Генетикалық және клеткалық инженерия тәсілдері
Жоспары:
1Генетикалық инженерияның міндеттері
2клеткалық инженерия
3Генетикалық инженерия
4Генетикалық және клеткалық инженерияның практикалық маңызы.
Дәрістің мақсаты:Генетикалық инженерия туралы түсінік алу.
Генетикалық инженерияның міндеттері
Генетикалық инженерияның көздеген мақсаты мен зерттеу әдістеріне байланысты мынандай деңгейлерін ажыратуға болады:1)малекулалық( геннің құрамды бөліктерің зерттеу,2)гендік 3)хромосомалық 4)клеткалық5) ұлпалық6) организмдің 7)популациялық
Бұл саладағы алғашқы зерттеулердің қатарына Н.П.Дубининнің (1934ж) дрозофила шыбындарының хромосом санын өзгнрту бағытында жүргізген жұмыстары жатады.Ол рентген сәулелерімен әсер ету жолымен хромосома санын өзгертуге болаындығын анықтады.
1956 Е.Сирс рентген сәулесінің көмегімен жабайы эгилопс өсімдігі жапырағының сары дақ ауруына төзімділігін анықтайтын генді жұмсақ бидайдың хромосомасына апарып салған.Жұмсақ бидайдың сары дақ ауруына төзімді формасы алынған.
1971 жылыВ.А.Струников ген және хромосом деңгейіндегі генетикалық инженерия әдістерін жібек құртында пайдаланады.
Клетка деңгейінде жүргізілетін жұмыстардың ішінде сомалық клеткаларды гибридзациялау әдісі кеңінен қолданылады.1960 жылы Ж.Барский жануарлардың сомалық клеткаларының бір бірімен қосылып, екі клеткадағы генетикалық информацияны біріктіруге қабілетті екендігін көрсетті.
Сомалық клеткаларды гибредтеу өсімдіктерге де жүргізіледі. Темекі , сәбіз, томат сияқты өсімдіктердің жеке протопластарынан клетка қабықшасы регенерациланып, калус түзілгеннен кейін, тұтас өсімдік қалыптасқандығы анықталды.
К.Грежник (1973ж) көп жыл бойы лабараторияда жүргізген зерттеулердің нәтежесінде сомалық клеткалардың түр ішіндік және түр аралық гибредтерін алған.
Генетикалық инженерия
Ген инженерияның қалыптасуы 1972 басталады. Осы жылы америка оқымыстысы П.Берг тұңғыш рет маймылдың онкогенді вирусының , бактериофагтың және E.Coli бактериясы геномдарының түрлі бөліктерін біріктіру арқылы рекомбинантты ДНК алды.
Активті гибридті ДНК малекуласын ең бірінші болып 1974 жылы С.Коян, Д.Хеленский, Г.Бойер алды. Олар құрамында әртүрлі бактериялардың, бақа ооцитінің теңіз кірінісі, тышұан митохондриясы ДНК-ларының фрагменттері енетін «химерлі» плазмидтері құрастырып шығарады. Оларды трансформация жолымен бактерия клеткасына енгізу арқылы онда жоғары сатыдағы организмдер ДНК-сына тән транскрипция құбылысын жүргізуге мүмкіндік туды.
Ген инженериясының теориялық негізінде генетикалық кодтың универсалдығы жатады.Бір ғана кодтың (триплеттің) барлық организмдердің адам, жануар, өсімдік, бактериялардың белок молекулаларының құрамына енетін амин қышқылдарын бақылай алатындығына байланысты ДНК малекуласының кез келген бөлігін басқа клеткаға апарып салу, яғни малекулалық деңгейде гибредтеу теориялық тұрғыдан алғанда мүмкін болып есептеледі. Генді бөтен клеткаға апарып салу (трансгеноз) және геннің өзін ДНҚ құрамынан бөліп алу немесе қолдан синтездеу күрделі жұмыстар. Ал организмге сырттан енгізілген геннің жаңа генетикалық аппарат құрамына қосылып, қызмет атқаруы одан да күрделірек. Бірақта бұл салада қол жеткен табыстар баршылық.Ж асанды ортада балапан ұрығының клеткасы өсірілген. Белгілі бір уақытта оған жаңа синтезделген ДНК жіпшелерінің құрамына енетін бромдезоксиуридин қосылған. Осылайша таңбаланған жаңа синтезделген ДНК -ны бұрынғы ескі ДНК-дан оңай ажыратуға болады.Сонымен қатар осындай ортаға тышқан клеткасынан бөлініп алынғын тритимин (H3)таңбаланған ДНК қосылған.
Содан біраз уақыттан соң клетка көбейгеннен кейін, оның құрамындағы генетикалық материалды алып қарағанда тышқанның ДНК-сы мен балапан ДНҚ-сының араласып кеткендігі байқалған.
ДНҚ құрамынан жеке генді бөліп алу жұмысы тұңғыш ре 1969 жылы Дж. Беквитің лабораториясында жүргізілген. Ол E.Coli бактериясынан лактозалық оперон тобына жататын гендерді бөліп алды.
Гендік инженерия әдістерімен рекомбинантты ДНК құрамына енетін жекеленген гендерді мынадай әдістермен алуға болады:
1)табиғи ортадан (клетка, организм) тікелей бөліп алу, 2) химиялық жолмен синтездеу 3)белгілі бір генге сәйкес келетін иРНҚ-ның көшірмесін алу.
Бірінші әдіс ген инженерияның дамуының алғашқы кезеңінде қолданыла бастады.Белгілі организмнің ДНК-сын түгелімен әр түрлі рестрикатазалармен үзіп, әр түрлі фрагменттер алады. Сонан соң оны клетка ішіне «арқалап» кіргізе алатын сақиналы (дөңгелек) плазмидалармен жалғайды. Ол үшін плазмиданы да рестриктазалармен үзіп, әр т үрлі фрагменттер алады. Сонан соң оны клетка ішіне «арқалап» кіргізе алатын сақиналы (дөңгелек) плазмидалармен жалғайды. Ол үшін плазмиданы да рестриктазалармен үзеді, оған плазмидалардың әрқайсысының құрамында бір немесе бірнеше бірнеше бөтен ДНҚ фрагменті (гені) болады.Одан кейін ол плазмидалары қайтадан бактерияға енгізеді. Осының нәтижесінде бактерия клеткасының әрқайсысында басқа организм генінің бір түрі болады. Осындай әр түрлі бөтен гендері бар бактерия клеткаларының жиынтығын немесе колекциясын «гендер банкі», кейде «гендер кітапханасы» деп атайды. Зерттеушілер ол банкіден керек уақыт ында қажет белоктың генін жаңадан тауып алады.Осындай гендер банкі қазір Ресейде, Батыс Европада және АҚШ-та жасалған.
Химиялық жолмен жасанды генді 1969 жылы Г.Корана синтезделген . Бірақ оған жалғасқан промотор тізбегі мен транскрипцияны аяқтайтын кодондар болмағандықтан, ол клетка ішінде ешбір қызмет көрсете алмады. Гендерді химиялық синездеуге нуклеин қышқылдарындағы нуклеотидтердің орналасу тәртібін анықтау әдісін тапқаннан кейін ғана мүмкіндік туды. Бұл әдістерді тапқан Д.Джиильберт пен Ф.Сэнгер.
Ғалымдар генді белоктың құрамындағы амин қышқылдарына қарап отырып синтездеуді де үйренді.(3нуклеотид -1кодон -1амин қышқылы деген заңдылық бойынша). Соның ішінде қолдан синтезделген ұзын ген адамның самототропин (өсу) гені, ол 584 нуклеотидтен тұрады. Оны бактериядағы басқа геннің промоторына жалғастырып, плазмида арқылы бактерия клеткасына енгізді. Соның нәтежесінде бактерияның бір клеткасы 3 млн га дейін адам самотропин малекуласын жасай алатын болды. Адам инсулин де химиялық жолмен синтезделіп, әлгі айтылған жолмен бактерияға енгізілді.
1979 жылы біздің елімізде Ю.А.Овчинников пен М.Н.Колосовтың басшылығымен ферменттердің көмегімен химиялық жолмен адам мен жануалардың гармонның гендері –онкефалин және брадикинин синтезделеді. Химиялық ферментік синтездеу ұсақ гендерді алу үшін ген инженериясында кеңінен қолданылады.
1970жылы Г.темин Т.Мизутаин және Д.Балтимор кері транскриптаза (ревертраза) ферментін ашты.1972 жылы кейбір онкогенді (рак) вирустар кері транскриптаза ферментінің көмегімен РНҚ ның үлгісінен ДНҚ синтездейтіні ашылды. Одан әрі жүргізілген зерт теулер ДНҚ көшірмесінің пайда болуы үшін онкогендік вирустардың ғана РНҚ сы емес, сондай ақ жасанды подирибонуклеотидтердің де үлгі бола алатындығын көрсетті.Бұл кез келген жеке гендерді (ДНҚ), олардың РНҚ көшірмелерін қолдана отырып ферменттік синтездеуге болатынына мүмкіншілік туғызады. Жасанды генді рак вирустарынан бөлініп алынған кері транскриптазаның организмнен алынған РНҚ ға қарап, оның тізбегіне сәйкес генді (ДНҚ ны) жасауға болады. Осы әдісімен самототропиннің гені жасалынып алынды.Ол үшін гипофиздің рак клеткаларынан бөлініп алынған самототропиннің иРНҚ сы пайдаланылды (иРНҚ рак клеткаларында өте көп синтезделеді екен)
ССРО да академик Ю.А.Овчинниковтың басшылығымен интерфон гені кері транскриптазха арқылы жасалып, ақырында ең көп мөлшерде интерфон жасайтын бактерия түрі алынд.
Ферментті синтездеп пробиркада РНҚ малекуласынан ДНҚ генінің, комплеменарлы тізбегән жазып алуды транскрипция дейді.Синтездеу үшін қолданылатын жүйе құрамында ДНК ға кіретін төрт нуклеотид, магний ионы, кері транскриптаза, оған сәйкес ДНК тізбегін синтездеп, сол ферменттің көмегімен ДНК ның екінші тізбегі синтезделеді. Осының нәтежесінде иРНҚ да синтезделген ген құрылымына ұқсас ген пайда болады.Осы әдіспен В.А.Энгельдерт басшылығымен «ревертаза» жобасы жасалған. Осы ферменттің көмегімен гендер синтезделеді. Бұл жобаны іске асыру үшін көптеген институттар , сондай ақ ғылым академиялары қатысты.
Генетикалық және клеткалық инженерияның практикалық маңызы
Малекулалық генетика мен генетикалық инженерияның қол жеткен табыстарының арқасында клеткаға сырттан жаңа генетикалық информация енгізу арқылы организмдердің алдын ала жобаланған тұқым қуалайтын бағдарламасын жасауға болады.Ондай информациянының қолдан синезделенуі мүмкін немесе табиғи генетикалық зат ретінде әртүрлі организмдерден бөлініп алынады.Нәтежесінде , эксперементтік әдіспен табиғи эволюциялық жолмен пайда бола алмайтын жаңа генетикалық жүйе жасалынады.
Генетикалық инженерия денсаулық сақтау, ауылшаруашылығы, биотехнология және микробиологиялық өндіріс салаларында маңызды рөл атқарады.Генетикалық инженерия әдістерін жетілдіру денсаулық сақтау мен ауыл шаруашылығы үшін қажетті гармондар, ферменттер және антибиотиктер синтездейтін микроорганизмдердің жаңа штамдардың алуға мүмкіндік туғызады.Жануарлар гендерін бактерияларға жіберудің де үлкен практикалық мәні бар.
Ауыл шаруашылығында, оның ішгінде егіншілікте өсімдіктердің атмосфералық азотты өздері жинақтап алуы басты проблема болып есептеледі. Азот белоктың құрамына кіретін негізгі компонент. Азотты, кейбір өсімдіктер , мысалы бұршақ тұқымдастар топырақ бактерияларымен селбесіп, өздері жинақтайды. Көпшілік өсімдіктерде, әсіресе, астық тұқымдастарда ондай қажет жоқ, сондықтан олар үшін миллиондаған тонна азотты тыңайтқыштар жұмсалады. Олар топыраққа 40 пайыздайы ғана сіңіп, қалғаны сумен шайылып кетеді. Екінші жағынан ол экологиялық тұрғыда тиімсіз болып есептеледі, себебі көп мөлшердегі нитратты қосылыстар тірі организмдерге зиян кел іреді. Соңғы кезде бұл фиксациялауға қабілеті жоқ пішен таяқшасына азот жинақтай алатын басқа бір бактерияның генін апарып салған , сонда ол азот фиксациялау (бекіту) қасиетіне ие болған. Осындай зерттеулердің негізінде аталмыш қасиеті астық тұқымдас өсімдіктерге беру мүмкіндігі бар екендігі анықталды.
Денсаулық сақтау саласында тұқым қуалайтын ауруларды емдеу үшін ген инженериясының үлкен маңызы бар. Қазіргі кезде ауру адамдардан зат алмасудың 1000 нан астам түрлі тұқым қуалайтын өзгерістер табылған. Соның салдарынан белгілі бір гендердің муацияға ұшырауына байланысты ферменттердің , гармондардың немесе белоктардың өзгеретіндігі анықталды.Ондай аурулардан мүлдем айығу үшін сол мутантты гендердңі жөндеу керек немесе өзгерген генді сау генмен алмастыру қажет. Галактоземия ферменті синтезделмейді. Соған байланысты адам организмі сүт қанының құрамына енетін галоктозаны сіңіре алмайды. Содан келіп галактоза бауыр мен мидың т.б органдардың клеткаларына жинақталады. Соның салдарынан көз көрмей қалады, бауырдың қызметі бұзылады, ақыл кемиді т.б күрделі өзгерістер пайда болады.
Бұл жағдайда адам клеткасына ондағы жетіспейтін функцияны қалпына келтіру үшін ген жіберу керек. 1971 жылы Меррил мен оның қызметтестері галоктоземия ауруы бойынша адам клеткасында жүргізілген генотерапияның сәтті аяқталғанын хабарлады. Олар галоктоземияны ауруы бойынша адам клеткасында жүргізілген генотропияның сәтті аяқталғанын хабарлады.Олар галактозаны сіңіруге қажетті ферменті болмаған. Оны қалыпқа келтіру үшін құрамында галактоза гені бар E.Coli бактериясының фагы алынған. Сондай бактерия генін адам клеткасына жібергенде ол галоктоза ферментін синтездеуге кіріскен. Австралия оқымыстысы Дой 1973 жылы томат өсімдігінің гаплойдты клеткаларын алған. Егер ондай ортаға ДНК сына жіберу жоғары сатыдағы организмдердің генетикалық материалының молекулалық құрылымын зерттеудің жаңа бір тәсілі болып есептеледі. Ген инженериясының әдістері гендердің қызметі мен құрылымдық ерекшеліктеріне тереңірек үңілуге мүмкіндік береді.Ген инженериясының әдістері гендердің қызметі мен құрылымдық ерекшеліктеріне тереңірек үңілуге мүмкіндік береді. Ген проблемасын зерттеудің мұндай деңгейі канцорегенездің имммунологиялық қасиетін, аллергияның практикалық медицинада маңызы бар басқа да құбылыстардың мәнін ашуда ерекше маңызы бар.
Бақылау сұрақтары:
- Генетикалық инженерияның теориялық және практикалық мәні?
- Генетикалық инженерия бойынша зерттеулер қандай деәгейде жүргізіледі?
- Геицина үшін гетикалық инженерияның медицина үшін қандай м аңызы бар?
- Ауыл шаруашылығын дамытуда генетикалық инженерия қандай роль атқарады?
- Генетикалық инженерия селекцияда не үшін қолданылады?
Пайдаланылатын әдебиеттер:
- Инге-Вечтомов С.ГюГенетика сосновами селекции М.Высшая 1989.
- Лобашев МюЕ.Ватти Кюе.Тихомирова М.МюГенетика с основами селекция М.просвещение 1979
- ватти К.В.Тихомирова М.М.Руководства к пратическим занятиям по генетике М.просвещение 1979 1972
- Лобашев М.Е.Генетика. Изд-во ЛГУ..Генетические 1969.
- Медведов Н.Н.Пратическая генетике М.Наука 1966.
- Мұкамбетжанов К.К Далабаев Б.А Өтешова Г.А Генетикадан практикалық сабақтар Алматы.Ғылым2004.
- Мұхамбетжанов К.Қ. Генетика. Алматы 2005
21 Дәріс
Дәрістің тақырыбы: Популяциялық генетика
Жоспары:
1.Популяция және оның генетикалық құрылымы.
2.Гарли-Вайнберг заңы.
Дәрістің мақсаты: Популяциялық генетика және эволюцияның генетикалық негіздері туралы мағлұмат алу.
Популяциялық генетика және оның генетикалық құрылымы.
Спетематиктердің айтуынша қазіргі кезде жануарлардың 2миллиондай, ал өсімдТіктердің 500 мыңдай түрі белгілі.Осыншама түрлердің пайда болуы және сақталуы тұқым қуалаушылық пен өзгергіштіктің заңдылықтарына байланысты екені сөзсіз.Тірі организмнің өзін қоршаған сыртқы ортаға бейімделуі барысында табиғи жолмен сұрыпталу арқылы жаңа түрлердің пайда болатындығы туралы тұңғыш рет түсіінік берген адам Ч.Дарвин.Ол эволюцияның механизмін құрайтын басты үш процестің, атап айтқанда тұқым қуалаушылықтың өзгергіштіктің және сұрыптаудың болатындығын анықтады.Бұл үш процесті К.А.Тимирязев эволюцияның шешуші факторлары деп атаған.
Түрді белгілі бір ариялға тараған, шығу тегі бірдей, сыртқы орта жағдайларына бейімделуі де ұқсас организмдердің тарихи қалыптасқан жиынтығы деп қарастыруға болады.Бір түрге жататын организмдердің өзіне тән генотипі және фенотипі бар.Кейбір түрлердің таралу ариялы кең, оған бірнеше географиялық нәсілдер (раса) немесе түр тармақтар енеді, оларды политипті дейді.Ал енді біреулерінің ареалы шектеулі болып келеді де географиялық нәсілдер құрай алмайды, ондай түрлерді монотипті деп атайды.Түрді құрайтын особьтар өзгермейтін біртекті масса емес.Белгілі бір түрге жататын әрбәр организмнің сол түрге тән жалпы қасиеттемен қатар өзімдік жеке генотиптік ерекшеліктеріде болады.Түр организмдердің біртұтас жиынтығы бола отырып, жекелеген популяцияларға бөлінеді.
Популяция деп өмір сүру ортасы бір және оған бейімделу қабілеті де бірдей, бір-бірімен бодандасып ұрпақ бере алатын бір түрге жататын дарақтарды атайды.Қолдан сұрыптау жолымен алынатын жануарлар тұқымдарымен өсімдіктер сортарыда жекелеген популяцияға жатады.Популяцияның қалыптасу процесі және оның диномикасы микроэволюция болып есептеледі.
Популяцияның генетикалық құрылымын анықтау селекционерлерден басалады деуге болады.Себебі, олар қандай болмасын бір сорт не тұқым шығару үшін бодандастыратын ата-ана жұбын таңдап алумен қатар, бірнеше буын бойы олардың ұрпақтарын зерттейді.Бірақ популяцияны генетикалық тұрғыда зерттеудің ғылыми негізі тұқым қуалаушылықтың заңдылықтары, яғни Мендель заңдары ашылғаннан кеін салынды.
Популяция құрылымын ең алғаш статистикалық және генетикалық әдістер қолдана отырып зерттеген дат генетигі В.Иоганнсен.Оның 1903 жылы «Популяциядағы және таза линиялардағы тұқым қуалау» деген еңбегі жарық көрді.
Иоганнсен популяцияны зерттеу үшін объект ретінде өздігінен тозаңданатын өсімдіктерді алды, себебі олардан таза линияларды бөліп шығару оңай.Ол бұршақ тұқымдарының салмағын өлшеп, оған талдау жасаған.Сонда олардың салмағы 150-750 мг аралығында болған.Содан кейін салмағы 250-350 тұқымдары бір бөлек семірген.Олардан өсіп шыққан өсімдіктердің тұқымдары қайтадан өлшенген.Бір сортқа жататын бұршақтан іріктеліп алынған ауыр және жеңіл салмақты тұқымдардан өсіп шыққан өсімдіктер тұқымдарының да салмағы жөнінен айырмашылықтары болған, яғни ауыр салмақты тұқымдардан өсіп шыққан өсімдіктердің дәндері орта есеппен 518,7 мг ал жеңілдері 343,4 мг тартқан.Сонда бұл тәжірибенің нәтижесі бұршаұтың сорт-популяциясының генетикалық жағынан әр текті особьтардан тұратындығын және оның әрқайсысы таза линиялардың бастамасы бола алатындығын көрсетеді.Өздігінен ұрықтану жағдайында әрьір жеке организм жаңа түр тармағының немесе түрдің және сол сияқты сорт немесе тұқымның шығар көзі бола алады.Мысалы, бидайдың сортының популяция ішінен сұрыпталып алынған бір ғана дәннен шығуы мүмкін.Айқас тозаңданатын организмдердің популяциясы генотипте әр түрлі дарақтардың еркін бодандасуы негізінде қалыптасады.Ондай популяцияны панмексиялық популяция деп атайды.Айқас тозаңданудың өздігінен тозаңданудан айырмашылығы сұрыптау және сұрыпталған формаларды бодандастыру жолымен бастапқы популяциядан өзгеше белгілері.
Панмексиялық популяцияның генетикалық ерекшеліктерін білудің бір жолы-жекелеген гендер бойынша гомозиготалы және гетерозиготалы дарақтардың таралу сипатын зерттеу.Мысалы, қандай болмасын бір популяцияда бір геннің әр түрлі аллельдеу бойынша, айталық, АА және аа гомозиготалы формаларды сан жағынан бірдей десек мұндай панмексиялық популяция тең мөлшерде А және а гендері бар аталық және аналық гометалары түзе алады.Осындай гендерді алып жүретін дарақтар бодандасқанда мынандай комбинация шығады:
| 0,5 A | 0,5 a | |
| 0,5 A | 0,25 AA | 0,25Aa |
| 0,5 a | 0,25 Aa | 0,25 aa |
Сонда  де доминатты гомозиготаның пайда болу жиілігі – 0,25-ке, гетерозигота – 0,5-ке,реиссивті гомозигота – 0,25ке тең.Ал келесі буында гамета түзілгенде қайтадан доминанатты аллелі бар гаметаның мөлшері 0,5 ке тең болады, себебі 0,25 гетерзигота Аа-дан келеді.ал енді рецессивті (а) аллелі бар гаметаның мөлшері де 0,5-ке тең.Себебі 0,25 рецессивті гомозигота аа-дан, ал 0,25 гетерзиготадан Аа беріледі.Сөйтіп, бұл мысалдан еркін будандасу жағдайында әр буын сайын доминатты және рецессивті гендері бар гаметалардың мөлшері бірдей деңгейде, яғни 0,5 A және 0,5 а болып отыратындығы байқалады.
де доминатты гомозиготаның пайда болу жиілігі – 0,25-ке, гетерозигота – 0,5-ке,реиссивті гомозигота – 0,25ке тең.Ал келесі буында гамета түзілгенде қайтадан доминанатты аллелі бар гаметаның мөлшері 0,5 ке тең болады, себебі 0,25 гетерзигота Аа-дан келеді.ал енді рецессивті (а) аллелі бар гаметаның мөлшері де 0,5-ке тең.Себебі 0,25 рецессивті гомозигота аа-дан, ал 0,25 гетерзиготадан Аа беріледі.Сөйтіп, бұл мысалдан еркін будандасу жағдайында әр буын сайын доминатты және рецессивті гендері бар гаметалардың мөлшері бірдей деңгейде, яғни 0,5 A және 0,5 а болып отыратындығы байқалады.
Бірақ кейде популациядағы гомозиготалар санынан тең болмайтындығы да кездеседі.Мысалы, қара бидайда сабағынан түктілігін анықтайды. А және түгінің жоқтығын анықтайтын а аллельдері бар.Егер қара бидайдың бір популациясында түгі бар өсімдіктер түгі жоқ өсімдіктерге қарағанда төрт есе көп болып келсе ( 4 АА : Laa), бұл жағдайда гаметалардың арақатынасы 0,5 A : 0,5 a емес 0,8 A : 0,2 a болып келеді.Будандасудың нәтижесінде олардан мынандай комбинация түзеді:
| 0,8 A | 0,2 a | |
| 0,8 A | 0,64 AA | 0,16Aa |
| 0,2 a | 0,16Aa | 0,04aa |
Сөйтіп, әр жүз өсімдіктің 96-сы түкті ( 64-і гомозигота, 32-і популацияда гетерозигота), тек
4-уі ғана түксіз болып шығады.
Бұл жағдайда келесі буындарда да «а» аллелі бар гаметалардың мөлшері 0,2 (0,04-гомозигота аа- дан, 0,16 – гетерзигота Аа-дан келеді).ал «А» аллелі бар гаметалардың мөлшері 0,8 болады ( 0,64 гомозигота АА – дан, 0,16 гетерозигота Аа-дан).
Бұл популацияда да бірнеше буын бойы гендердің белгілі бір ара қатынасы, яғни 0,8:0,2 сақталады.
Харди мен Вайнберг заңы
1906 жылы ағылшын математигі Харди мен дәрігері Вайнберг бір-біріне тәуелсіз түрде популациядағы генотип пен фенотиптің таралуын көрсететін формула ұсынды.ол кейіннен Харди – Вайнберг заңы деп аталады.Бұл формуланы құрастырғанда олар аллельдердің мөлшері өзгермейтін жағдайда популациядағы доминантты және рецессивті белгілері бар дарақтардың белгілі бір арақатынаста болатындығын негізге алды.
Егер гаметадағы бір аллельдің,айтайық, А аллелінің мөлшерін Р деп белгілесек,онда екінші алдель а-ның мөлшері q болады.Осыдан келіп олардың ұрпақтарында мынадай арақатынас шығады.
| P A | q a | |
| pA | 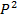 AA AA
| 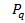 Aa Aa
|
| qa | Aa
|  aa aa
|
Бұл адгандарды жинақтағанда Харди-Вайнберг формуласы шығады:
AA : 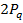 Aa:
Aa: 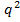 aa
aa
Мысалы, ірі қараның бір популациясында мүйізділері 25% немесе 0,25 болып, ал тоқалдары 75% немесе 0,75 болып келсе,тоқалдық белгіні доминантты ген «А», ал мүйізділігін рецессивті ген «а» анықтайды.Генотип «аа» ның мөлшері бұл жағдайда 0,25 болғандықтан а аллелінің мөлшері а= 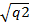 ,
, 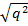 =
= 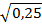 =0,05, a=0,05.
=0,05, a=0,05.
Доминантты аллель А-нын мөлшері де осы формулаға сәйкес анықталды:
P=I-q=I-0,05=0,95
Сонда популациядағы доминантты генотип АА ның мөлшері
AA =0,9 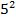 =90,25
=90,25
Рецессивті және доминантты гомозиготалы генотиптердің мөлшеріне қарап, популациядағы гетерозиготалардың мөлшерін анықтауға болады.
Aa=2 ·0,95·0,05=9,5
Харди мен Вайнберг формуласын пайдалана отырып, белгілі бір фенотиптің мөлшерін есепке алудың негізінде популациядағы генотиптің таралу сипаты анықталды.Бірақ бұл формуланы қолданудың да белгілі бір шегі бар.ол мынандай жағдайлар: егер аутосомдық гендердің бір жұбы есепке алынса,доминантты аллельдегі мутацияның рецессивті аллельге немесе керісінше ауысуы болмайтын жағдайда: әртүрлі генотипті дарақтардың тіршілік қабілеті және ұрпақ беруі бірдей болғанда.Табиғи популацияларда бұл қойылатын талаптардың бәрі бірдей бола бермеуі мүмкін.Сондықтан Харди – Вайнберг формуласын барлық жағдайда бірдей қолдана беруге болмайды.Бұл заңдылықтың үлкен теориялық та, практикалық та мәні бар.бұл формуламен адамдар арасындағы кейбір белгілердің таралу жиілігі анықталады.Мысалы,қан топтарын анықтайтын гендердің мөлшерін есептеуге болады.
Өздігінен ұрықтанған организмдердің популациясы үшін Харди- Вайнберг формуласы қолданылмайды.Өздігінен ұрықтанған кезде гетерозиготалы гендер гомозиготалы жағдайға көшеді.
Бақылау сұрақтары:
- Популациялық генетика ұғымын бірінші рет енгізген ғалым?
- Панмиксиялық популацияның генетикалық ерекшелігін ата?
- Айқас тозаңданатын организмдердің популациясы қандай дарақтардан тұрады?
-
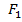 – де доминатты гомозиготаның пайда болу жиілігі?
– де доминатты гомозиготаның пайда болу жиілігі? -
 - де гетерозиготаның пайда болу жиілігі?
- де гетерозиготаның пайда болу жиілігі? -
 мен Вайнберг формуласы қалай болады?
мен Вайнберг формуласы қалай болады?
Пайдаланылған әдебиеттер:
- Инге-Вечтомов С.Г.Генетика сосновами селекция ,М.Высшая школа 1989.
- Лобашев М.Е.Ватти К.Е.Тихомирова М.М. Генетика с основами селекция М.Просвещение 1979
- Ватти К.В.Тихомирова М.М.Руководства к практическим занятиям но генетика. М.Просвещение 19791972.
- Лобашев М.Е.Генетика. Изд-во ЛГУ.. Генетические 1969.
Медведев Н.Н.Пратическая генетика
22 дәріс
Дәрістің тақырыбы: Популациялардың генетикалық динамикалық факторлары
Жоспары:
- Мутациялық процесс
- Сұрыптау
- Популацияның мөл
- Изоляция
Мутациялық процесс
Гендердің популяцияда тұрақты түрде қайталанып тұруы үшін олар мутацияға ұшырамауы керек. Бірақ ондай өзгеріс үнемі болып отырады.Әрбір жеке ген мутацияға сирек ұшырағанмен, генотиптегі гендердің саны көп болғандықтан, түрлі мутациялардың жалпы мөлшері де едәуір көп болып келеді.
Популациядағы генотиптері геерозиготалы организмдер түрлі мутацияларға бай болады.Мысалы, жүгерінің әртүрлі сорттарында «жасыл өскінді» мутация бойынша гетерозиготалы өсімдіктер 34-66 пайызға дейін кездеседі.
Әрбір буын сайын генофонд (тектік көр) жаңа мутациялардың жиынтығымен толықтырылып отырады.ол процесті мутациялық қысым деп атайды.Қандай болмасын жаңа пайда болған мутация түрге тән генотиптің тұтастығында өзгеріс тудырады.Сол себепті генотип системасында сұрыптаудан өтпеген мутация жеке даму барасында организм функцияларының тарихи қалыптасқан корреляциясын (бір біріне сәйкестілігінін) бұзады. Сондықтан да мутация алғашқыда көбінесе зиянды болып келеді.
Сұрыптау
Сұрыптау деп генотиптің организмнің сыртқы орта жағдайларына мейлінше көп бейімделуін қамтамасыз ету процесін айтады. Организмнін тіршілік ету және ұрпақ қалдыру мүмкіндігі оның ортаға бейімделу дәрежесіне байланысты.Организмде бейімделу реакцияларының нормасы неғұрлым жоғары болса, оның популяция құрамында сақталып, дами беру мүмкіндігі соғұрлым мол болады.
Дарвиннің табиғи сұрыпталу туралы теориясы артта қалатын ұрпақ мөлшері олардың ата аналардың саналық ерекшеліктеріне байланысты деген принципке негізделген.Егер белгілі бір генотипі бар организмдер сұрыптаудың нәтижесінде жарамсыз болып қалса, онда популяциядағы соған тиісті геннің мөлшері азаяды,яғни сұрыптау тиімсіз гендердің таралуын бөгейді.
Әрбір буын сайын популяциядағы зияны көбірек мутацияның концентрациясы зияны азырағына қарағанда тез төмендеп отырады.
Доминатты және рецессивті аллельдердің популяциядан шығып кету жылдамдығы да әртүрлі болады.Доминантты летальді гені бар организмдер сұрыпталу арқылы бірінші буынның өзінде шетке шығып қалады.Басқаша айтқанда доминанты ген әрбір буын сайын үнемі сұрыптаудың бақылауында болады.Рецессивті мутация доминанты қарағанда популяция құрамында ұзақ сақтала алады.Сондықтан да рецессивті гендердің сұрыпталуының доминантты гендерге қарағанда нәтижесі төмен болады.
Сөйтіп, гендердің сұрыпталуы түрлерінің дивергенциясына шешуші роль атқарады.
Популяцияның мөлшері
Гендер концентрациясы популяциясының мөлшеріне байланысты болады.
Популяцияның мөлшері неғұрлым аз болса, гетерозиготалы дарақтардың бір бірімен будандасып, рецессивті гомозиготалы ұрпақ беруінің мүмкіндігі соғұрлым мол болады.Керісінше популяцияның мөлшері көп болса рецессивті гомозиготаның болу мүмкіндігі азаяды.Ал санды популяцияда сұрыпталу нәтижесінде пайдасыз гендер кеміп, гендер артып отырады.
1931 жылы Н.П. Дубинин сан жағынан популяцияның өзінде,кейде кейбір кездейсок факторлардың әсерінен, гендер концентрациясының өзгертетіндігін көрсетті.Кейіннен популяцияның саны артқан сайын ол гендердің мөлшері де арта түседі.Бұл құбылысты генетикалық – автоматты процесс немесе гендердің дрейфі деп аталады.