АЛМАСУЫ
Жыныстық жолмен көбейетін барлық организмдерде жеке фазалардың кезектесуі байқалады, мүндай фазалардың алмасуында хромосомалар саны бір. есе болатын гаплофазалар, хромосомалар саны екі есе болатын диплофазалар алмасып отырады. Бұл фазалардың ұзақтығы және дамуы әр турлі организмдерде түрліше болады. Көптеген жануарларда гаплофаза редукцияланып қысқарған жәие ол жыныс клеткасы түрінде ғана болады (қүрттар, насекомдар, балықтар, қосмекенділер, құстар, сүтқоректілер т. б.).Ал нейроспора (Ыеигозрога сгазза), аспергилла (Азрег§і1-Іиз пісіиіапз) сияқты төменгі сатылы өсімдіктерде керісінше диплофаза зигота стадиясына дейін редукцияланған, олар пайда болысымен мейоздан өтеді де гаплоидты споралар түзеді, одан тіршілік циклінің негізін құрайтын гаплофаза өсіп-жетіледі. Кейбір балдырларда гаплофаза мен диплофаза шамамен бірдей жетілген. Мүктерде гаплофаза күштірек жетілген бола-ды (диплофаза зиготадан өсіп-жетілетін спорангиямен шектелген), ал папоротниктерде диплофаза басым болады (гаплофаза ескінше түрінде болады). Гүлді өсімдіктер диплоидты, оларда гаплофаза редукциялаыған (микроспоралар мен ұрық қалтасы).
Тіршілік цикліндегі фазаларды білудің көптеген генетикалық қүбылыстарды зерттеуде маңызы зор (II, IV бөлімдерді қараңыз).Сонымен, жынысты көбею кезінде екі гамета косылыи, зигота түзілгеніне карамастан бірнеше үрпақ бойы әр түрдің езіне тән тұрақты хромосомалар саны сақталады. Бүл жағдай гаметалар түзілуі хромосомалар санының екі есе азаюын қамтамасыз ететін арнаулы механизмнің болуына байланысты екен. Мүндай механизм мейоз болып табылады. Ал ұрықтану хромосома санын диплоидты қалпына келтіретін механизм қызметін атқарады.
Жынысты көбею процесінде хромосомалар санының тұрақтылығы ғана сақталып қоймайды, сонымен қатар жиынтықтың тұтас кариотиптің түрақтылығы сақталады. Бүл гомологиялық хромосомалардың мейоздағы коньюгациясына, одан кейін олардың бөлініп полюстерге тарауы нәтижесінде бір гомолоттың бір полюске, ал екінші гомологтың екінші полюске баруьша байланысты. Бұл түрғыдан қарағанда жыныстық көбею мен жыныссыз көбею үқсас.
Егерде зигота кариотппін хромосомалардың саны мен құрылысы түрғысынан ғана қарастырмай, сонымен қатар хромосомалардың сапасын (шығу тегін) ескерсек, онда гаметалар түзілуі процесінде гаметалар қүрамына шешелерінің де, әкелерініц де хромосомалары кіреді. Бүны мейозда бөлініп полюстерге таралуда гомологиялық хромосомалардың жеке жүптарының өзара тәуелсіздігі қамтамасыз етеді. Егер I профазадағы әкелері мен шешелерінің гомологиялық хромосомаларынын өзара учаскелер алмасатынын еске түсірсек, онда мейоздан кейінгі гаметада рекомбинантты хромосомалар пайда болады, яғни ондай хромосомаларда әкелерінің де және шешелерінін, де учаскелері — локустар болады.
Сонымеи, жынысты көбею процесінің жыныссыз көбеюден айырмасы — пайда болатын гаметалар сапа жағынан өзара да, бастапқы клеткаға да ұқсас болмайды. Ал ұрпақ алмасқан кезде ұрықтану процесі көптеген жаңа формалардың түзілуіне себеп болады.Бөлімнің басында мынадай сұрақтар қойылған: ұрпақтың ата-анасына ұқсастығын қамтамасыз ететін материалдық негіз не болып табылады? Клеткада тұқым қуалаушы информацияны алып журетін не? Сірә, қандай да бір клетка компоненті тұқым қуалаушылықтың материалдық негізі болу үшін ол мынадай бірқатар шарттарға сәйкес болуы тиіс: 1) клеткада зат алмасуға байланысты функцияны атқаруы тиіс; 2) өзі өндіретін қабілеті болуы керек және 3) ол жаңадан түзілетін жас клеткалар арасында теңдей болып бөлінуі қажет.
Алдағы қарастырған тараулардан осындай шарттармен хромосоманың қанағаттандырылатыны белгілі. Ол клеткадағы зат алмасуды бақылайды (толығырақ 17, 22 тарауларды қараңыз), хромосома репликациялана алатыны және көбеюдің кез келген әдісіиде, жас клетхалар арасында ол теңдей болып екіге бөлінеді.
Жоғарыда қойылған алғашқы екі шартқа, хромосомалардан өзге, пластидтер імен митохондриялар сияқты цитоплазмалык құрылымдар да сәйкес келеді. Шындығында тұқым қуалаушы информацияны алып жүруі мүмкіндігі жөнінде кейбір күдіктену бар. Қлеткалардың бөлінуінде олардың таралуы кездейсоқ болады. Бұл күдіктің оншама негізі жоқ, ейткені олардың клеткадағы саны өте көп және олардың бәрі бірдей қызмет атқара-ды, яғни олар бірін-бірі алмастыра алады.
Клетканың жеке құрылымының түұқым қуалаудағы ролін қарастырған кезде бұл мәселеге біз тағы да тоқталамыз.
II бөлім
Белгілердің тұқым қуалау заңдылықтары және тұқым
қуалаушылық принциптері
Тұқым қуалаушылықтың материалдық негіздерін алғашын-да бір-біріне
тәуелсіз цитология және генетика зерттеді.
Цитология клетка бөлінуінің негізгі заңдылықтарын және ұрпақтарының ұдайы өзін-өзі ендіруі кезінде жеке құрылымдарының рөлін анықтады. Генетика белгілердің берілу ерекшеліктерін зерттеді. Ал генетикада белгілердің тұқым қуалау заңдылықтарын анықтауға мүмкіндік беретін әдіс ашылғанға дей-ін, тұқым қуалаудың материалдық негізін білу мақсатымен цитология мен генетика мәліметтерін байланыстыруға болмайтын еді.
return false">ссылка скрыта4-тарау. ГИБРИДОЛОГИЯЛЫҚ ӘДІС
Тұқым қуалау заңдылықтарын зерттеу әдісін 1865 жылы Грегор Мендель анықтады.
1. ГИБРИДОЛОГИЯЛЫҚ ӘДІСТІҢ ЕРЕКШЕЛІКТЕРІ
Мендельге дейінгі ғалымдар, жүздеген жыл бойы будан ұрпақтардан белгілердің барлық жиынының тұқым қуалауын тұтас алып зерттеді. Ал Г. "Мендель тұқым қуалаудың жаңа принциптерін зерттеудің негізін салды.
Мендель әдісінің бірінші ерекшелігі бірнеше ұрпақ бойы коистаытты формалар алудан тұрады, бұдан кейін осы формаларды шағылыстырады.
Мендель әдісінің екінші ерекшелігі бір түрге жататын өсімдікті (бүршақ Різит заііуит) шағылыстырып, олардан алынған үрпақтарда жеке жүп белгілердің түқым қуалауына анализ жасау болды. Шығылыстырылатын әсімдіктердің бір, екі және үш жұп айқын көрінетін, альтернативтік белгілері болуы тиіс. Мысалы, гүлдері — қара қошқыл және ақ, тұқымының сыртқы пішіні бұдыр-бүдыр және тегіс т. б. Мұнда әр үрпақ бойынша осындай альтернативті белгілер жүбы жеке ескеріліп есептелді, бүл кезде өзара шағылыстырылған өсімдіктердің басқа айырмашылықтары ескерілмеді.
Бұл әдістің үшінші ерекшелігі — альтернативті белгілердің жеке жұптары бойынша ажыратылатын буданды өсімдіктерге бірнеше ұрпақ бойы сандық есептеулер жүргізіледі.
Мендель әдісінің төртінші ерекшелігі әр будан өсімдіктен алынған урпақ жеке қаралып оған анализ жасалды.
Зерттеудің аталған қарапайым әдістерінің өзі тұкым қуалаудың жаңа гибридологиялық әдісіне негіз болды. Ол тұқым қуалаушылық пен өзгергіштікті зерттеудің бүкіл бір дәуіріне есік ашты. Тұқым қуалаушылықты зерттеудің генетикалық әдістерінің жиынтығы генетикалық анализ деп аталады.
Белгілердің тұқым қуалаушылығына анализ жасап, оны баяндау алдында «белгі», «касиет» деген түсініктерді анықтап, генетикада қабылданған кейбір қысқарған сөздер (символдар) жөнінде айту қажет. «Белгі» ксмесе «қасиет» деген үғым организмнің морфологиялық, физиологиялық немесе биохимиялық дискреттілігі, бірлігінің шартты белгісі ретінде пайдаланылады. Біз бір организмді екіншілерінен олардың өздеріне тән ерекше-ліктері бойынша ажыратамыз. Мысалы, ондай ерекшеліктер көк көз — қой көз, қара шаш — сары шаш, ұзын бой — аласа бой, қанның тобы А-В т. с. с. болады.
Белгі дегеніміз бір организмді екіншісінен ажыратуға болатын жеке сапасы.
2. ШАҒЫЛЫСТЫРУДЫ ЖАЗУ ЕРЕЖЕСІ
Жынысты көбеюде белгілі бір белгілердің тұқым қуалауына генетикалық анализ жасау үшін жынысы әр түрлі екі организмді шағылыстыру қажет. Генетіікада шағылыстыруды (X) көбейту белгісімен көрсетеді. Шағылыстыру схемасын жазған кезде бірін-ші орынға ұрғашы жынысты организмді жазады. Ұрғашы жынысты белгісімен, еркек жынысты белгісімен көрсетеді.
Шағылыстыру үшін алынғаи ата-аналық организмдерді Р деген латын әрпімен белгілейді. Белгілері әр турлі екі организмді шағылыстыру иәтижесінде алынған үрпақ буданды деп, ал жеке особь бұдан деп аталады. Буданды ықшамдык үшіи Ғ әрпімен белгілейді, ол әріпке индекс қойылады, осы индекс бұданды үрпақтың реттік нөміріне сәйкес болады. Мысалы, бі-ріиші үрпақ Ғі болады; ал егер буданды организмдер өзара ша-ғылыстырылса, онда екінші ұрпақ Ғ2, үшінші үрпақ Ғ3 дел т. с. с. белгіленеді.
5-тарау. МОНОГИБРИДТІ ШАҒЫЛЫСТЫРУДАҒЫ
ТҰҚЫМ ҚУАЛАУ
Бастапқы ата-аналық формаларда ескерілетін айырым белгілер неше жүп екеніне қарай шағылыстыру моногибридті, д-гибридті немесе полигибридті деп ажыратылады.
|
21сурег. Бұршақ гүлінің қара қошқыл және ақ түсі-нін. тұқым қуалауы:Л— гүлдің ңара ңошңыл түсінің факторы; о— гүлдің аң түсінің факторы.
I. МОНОГИБРИДТІ ШАҒЫЛЫСТЫРУ
Моногибридті шағылыстыру деп ата-аналық формаларын кері альтернативті бір жұп белгі бойынша ерекшеленуін айтады. Мысалы, аталық өсімдіктің гүлі қара қошқыл, ал аналық өсімдіктікі — ақ, немесе керісінше болуы тиіс.
Шағылыстыру алдында әр ата-ана белгілерінің бірнеше ұрпақ бойы константты екенін, яғни өздігінен тозаңданғанда нмесе жақын туыстық шағылыстырғанда осындай әр белгі тұрақты тұқым қуалайтынын, ал өте көрнекті альтернативті белгілердің ұрпақта көрінбейтініне көз жеткізу керек.
Доминанттык, (басым болу).Будандардың біріншіұрпағының біркелкілік заңы. Бұршақ гүлі бояуының тұқым қуалауын моногибридті. шағылыстыру нәтижесінен қарастырып көрейік. Мендель де езінің нақтылы тәжірибесін осы дақылмен істеді (21-сурет). Егер аналық өсімдіктің гүлі қара қошқыл, ал аталық есімдіктікі ақ түсті болса, онда барлық будан өсімдіктер гүлінің түсі бірінші үрпақта Ғ\ қара қошқыл болады, гүлдің ақ түсі керінбейді.
Сонымен, ата-ананың біріне-бірі кері, альтернативті белгілердің біреуі ғана Ғх буданда жарыққа шығады, екінші белгі көрінбей қалады.
Будандардың бірінші ұрпағында ата-ананың біреуінің ғана белгісінің басым болуын Мендель доминанттыц деп атады. Бұданда көріне отырып, екінші альтернативті белгіні тұншықты-рушы белгіні доминантты деп, тұншыққан белгіні рецессывті деп атады. Осы құбылыс өсімдікке, жануарларға, адамға да универсал екені анықталды, сондықтан ол доминанттық ереже деп аталады.
Будандардың бірінші үрпағы бірдей, бір пішінді болатын-дықтан, бұл құбылыс Менделдің бідінші заңы деп, яки біріншІ Үрпақ будандарының біркелкілік заңы деп аталатын болды.
Ажырау заңьғ. Егер бірінші ұрпақ будандары Ғх өздігінен тозаңданса, онда к§лесі яғни Ғ2 ұрпағында ата-ана екеуінің де белгілері — қара қошқыл гүлдер де, ақ гүлдер де пайда болады. Екінші үрпақта ата-ананың екеуінің де белгілерінің (до-минант және рецессив белгілердің) көріну заңдылығы ажырау деп аталады. Ажырау кездейсоқ болмайды, ол белгілі бір саидық заңдылыққа бағынады. Атап айтқанда, екінші үрпақта Ғ2 алынған барлық өсімдіктердің 3/4 бөлігінің гүлі қара қошқыл түсті болады да, '/4 бөлігінің ғана түсі ақ болады. Доминант-ты өсімдіктер саны мен рецессивті белгісі бар өсімдіктер саны-ның аракатынасы 3 : 1 болады. Сонымен, буданның бірінші үр-пағтында рецессивті белгі мүлде жойылып кеткен жок, тек түн-шыктырылып қалған, ол екінші ұрпақта кайтадан көрінді.
Ғ2 кезінде будандардың доминанттық пен рецессивті белгілердің белгілі бір сандық қатынас түрінде ажырауы ажырау заңы немесе Мендельдің екіңші' задыдеп аталады.
Тұқым қуалаушы факторлар. Егер екінші ұрпақ өсімдіктері Ғ2 өздігінен тозаңданса, онда ақ гүлді (рецессивті белгі) есімдіктер келесі үрпақтарда — Ғ3 және Ғ4 т. с. с. — ақ гүлді өсімдік береді. Ал қара қошқыл гүлді өсімдіктер (доминантты белгілер) басқаша қасиет көрсетеді. Өздігінен тозанданған ка-ра кошқыл гүлді өсімдіктердің 1/3 белігі ғана Ғ3-те және одан кейінгі үрпақтарда қара қошқыл гүлді өсімдіктер береді де, қалған 2/3 бөлігі Ғ2 кезіндегідей кайта ажырайтын болады, яғни әрбір 3 қара қошқыл гүлді өсімдікке 1 ақ гүлді өсімдік келетін болады.
Сонымен, буданның бірінші үрпағында ата-аналардың біріне-бірі кері белгілерінід біреуі ғана болады, ал екінші рецессивті белгі көрінбейді, бірақ жасырынған белгі келесі, екінші ұрпақ өсімдіктерінің 1/4 бөлігінде жарыққа шығады. Осы фактілерден Мендель мынадай қорытынды жасады: түқым қуалай-тын белгілердің өздері емес, тұқым қуалау бастамасы, оларды анықтаушы факторлар. Мүндай жағдайда будандардың Ғл гүлдің ақ түсті белгісінің жоғалып кетуін және оның кейіннен екінші ұрпақ Ғ2 өсімдіктерінің бір бөлігінде пайда болуын толық түсіндіруге болады. Мүны былай деп топшылау керек: Ғ\ өсімдіктерінде ақ түстің жасырын бастамасы болған және ары қарай ұрпаққа (Ғ2) берілген.
Бұл бастамалар тұрақты және ұрпақтан ұрпаққа өзгеріссіз түрде беріліп отырады. Белгілі бір белгілердің дамуын анықтайтын тұқым қуалаушы факторлар ген деп аталды. «Ген» деген термин организмнің жеке бір белгісінің немесе қасиетінің дамып жетілуін анықтаушы тұқым қуалау бірлігі деп түсіну қажет. Тұқым қуалаушылық құбылысын және тұқым қуалау механизмін талдап, оларға анализ жасау тереңдеген сайын, ген табиғаты жөніндегі үғым да кеңейе береді.
Аллелизм.Мендель тұқым қуалаушы бастамаларды латын алфавитінің әріптерімен белгілеуді үсынды: доминантты бастаманы үлкен әріппен (мәселен, Л), ал рецессивті бастаманы соның кіші әрпімен (а) белгілеуді ұсынды. Бұл бастамалар әр организмде жұп болады, ейткені олардың біреуі шешесінен, екіңшісі әкесінен келеді.
Альтернативті белгілерді анықтаушы жұп генді аллеломорфты жұп деп атайды, ал жұптық құбылыстың өзін аллел-морфизм немесе аллелизм деп атайды. Әр фактор яки ген екі күйде — А және а болады, сондықтан олар бір жүп түзеді, ал жұптың әр мүшесі аллель деп аталады. Мысалы, бүршақ гү-лінің қара қошқыл және ақ түстері доминантты және рецессивті белгілері бір геннің екі аллеліне (доминантты және рецессивті) сәйкес келетін болады.
Генотип және фенотип.Ген аллельдерінің ерекшелігін ескере отырып бүршақ гүлдері бояуының тұқым қуалауын тағы да қарастырайық. Гүлдің қара қошқыл бояуының доминанттық аллелін А әрпімен, ал рецессивтік аллелін а әрпімен белгілей-міз. Осыған сәйкес қара қошқыл гүлді бастапқы өсімдіктің ал-лелі АА, ақ гүлді әсімдіктің аллелі аа болады. Белгілі бір ген-нің әр жүп аллельдерінің біреуі шешесінен, екіншісі әкесінен шыққан. Екі аллельдің біреуі ғана гаметаға беріледі, осыған байланысты ата-ананың әрқайсысы гаметаның бір сортын ға-на не А, немесе а береді. Аталық және аналық организмдердеы А және а аллельдерін алған бірінші ұрпақ будандарының гүл бояуы аналық өсімдіктікіндей (АА) қара қошқыл, бірақ оның түқым қуалаушы қасиеттерінің негізі өзгеше болады. Осыған орай белгілі бір қасиеті жарыққа шыққанның езінде, тұқым қуалаушы негіз әр түрлі (АА және Аа) болуы мүмкін. Сондықтан белгінің сыртқа шығуын немесе организмдер қасиеттері мен белгілерінің жиынтығын фенотшг деп атайды. Ал организм белгілерінің дамуын анықтайтын түқым қуалаушы бастамалардың жиынтығын генотип деп атайды. Сонымен, қарастырылып отырған мысалда аналық өсімдік пен Ғ\ буданңың фенотипі бірдей — екеуінің де гүлдерінің бояуы қара қошқыл, бірақ екеуінің генотипі әр түрлі — АА жәие Аа.
Бір геннің бірдей аллельдері бар организмдер, мысалы екі аллельдің екеуі де доминантты (АА) немесе екеуі де рецессивті (аа) болса, ондай организмдер гомозиготалы не гомозигота деп аталады. Ал егер организмде бір геннің екі түрлі аллелі болса және оның біреуі доминантты, екіншісі рецессивті (Аа) аллель болса, ондай организмдер гетерозиготалы не гетерезигота деп аталады. Сонымен, қарастырылған мысалда бастапқы ата-аналық өсімдіктер — гомозигота АА және аа, ал будандар — гетерозигота Аа болады. Гомозиготалы ата-аналық өсімдіктерден будан өсімдіктердің айырмасы — олар жұмыртқа клеткалары мен тозаңның екі сортын — А және а береді.
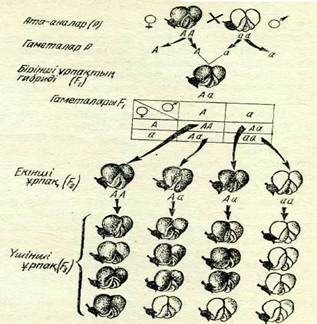
Пеннет торы.Әр түрлі гаметалар типтерінің өзара үйлесім-дерін есептеп шығаруды жеңілдету үшін ағылшын генетигі Р. Пеннет тор жасап, жазуды соған орналастыруды ұсынды, ол Пеннет торы деген атпен әдебиетке кірді (21-сурет). Бұл тор-да сол жақтағы вертикалда аналық, жоғары горизонтальда ата-лық гаметалар орналасады. Ал тордын, квадраттарына гамета-лардың үйлесу көрінісі жазылады. Бүл үйлесу зиготалардың генотиптеріне сәйкес болады.
Өздігінен тозаңданған жағдайда Ғ2 генотипі бойынша ІАА :2Аа-1аа қатынасында ажырайды. АА және Аа генотиптерінің фенотиптері бірдей, яғни гүл бояуы қара қошқыл бола-тындықтан Ғ2 белгілерінің фенотип бойынша ажырауы 3 қара қошқыл; 1 ақ гүлді қатынасында болады. Сонымен, белгілер-дің фенотип бойынша ажырауы генотип бойынша ажырауға сәйкес келмейді. Енді рецессивті аллельдері аа бар, ақ түсті гүлі бар гомозиготалы Ғ2 өсімдіктері өздігінен тозаңданғанда Ғъ кёзінде тек езі сияқты ақ гүлді өсімдіктер беретінін енді түсіндіруге болады. Мүндай өсімдіктер гаметалардың бір сортын ғана өндіреді. Сол сияқты қара қошқыл түсті гүлі бар есімдіктердің 1/3 белігі доминантты гомозиготалар (АА) болатын-дықтан, оларда ажырамайды, ал ондай қара қошқыл түсті есімдіктердің 2/3 бәлігі гетерозиготалы (Аа) болатындықтан және олардың генотипі Ғ\ буданыныкіндей болатындықтан, Ғ3 кезінде оларда ажырайды. Олардың белгілерінің ажырауы Ғ2 будандардікі сияқты, яғни 3 1 болады.
Гаметалар тазалығының ережесі. Моногибридті шағылыстыру кезіндегі белгілердің түқым қуалауына анализ жасай отырып, тағы бір маңызды қорытынды шығаруға болады. Егер Ғі буданында екі аллеломорфты белгілердің доминантты біреуі ғана көрінсе, онда Ғ2 кезінде бөлініп шығатын рецессивті белгі бастапқы ата-анадағыдай таза күйінде болуы, гетерозиготалар-да А және а аллельдерінің араласып кетпегенін көрсетеді. Нә-тижесінде, осындай гетерозигота түзген гаметалар «таза» бола-ды. Оның мағынасы мынадай: А гаметасы таза, рнда а алелльден қосылған ештеңе жок, сол сияқты а гаметасы А аллелінен таза болады. Гибрид гаметаларында альтернативті' белгілер жұбының аллельдерінің араласпау құбылысы гаметалар тазалығының ережесі деп аталды.
Түқым қуалаушы бастаманың салыстырмалы тұрақты болуы, будандастыру кезінде олардың араласпауы тұқым қуалаушылықтың дискреттілігін көрсетеді.

|
Дта-аналар (Р)
Гаметалар Р
БудандарҒ-і
Гаметалар Ғ^
2-сурет. Моногибридті будандастыру кезіндегі гомологиялық хромосомалар жұбының ерекше-лігін көрсететін схема:А—гүлдің қара ңошңыл түсінің факторы; а—гүлдің аң түсінің факторы.Будандар ҒМоногибридті ажыраудың цитологиялық механизмі. Ғ\ будандарынын, біркелкі болу себебі және будандардың Ғ2 кезінде ажырауы, сол сияқты факторлар жұбының табиғаты және гаметалар тазалығы Мендельге белгісіз болды. Өйткені Мендель өмір сүрген кезде клеткалардың бөлінуі де, гаметалардың тү-зілу процесі де, ұрықтануы да белгісіз еді. Алдыңғы тарауларда жыныс клеткаларының дамып жетілуімен және гаметалардың түзілуімен танысқаннан кейін, Мендель байқаған белгілердің ажырау құбылысын хромосомалар ерекшелігімен — олардың жұптығымен, мейозда гомологиялық хромосомалардын, ажыра-уымен және ұрықтану процесінде олардың қайта қосылып бірігуімен байланыстыруға болады.
22-суретті қарастырайық. Бүршақ өсімдігінің дене клеткасында бар болғаны — бір жүп қана гомологиялық хромосома-лар бар деп, ал гүлдің қара қошқыл түсін анықтайтын А әріп-мен белгіленген аллельддр* ата-аналық өсімдіктің әр хромосомасында бар деп алайық. Бүл жағдайда гүл бояуының доми-нантты белгілері бар томозиготалы өсімдіктердің дене клеткаларында гомологиялық хромосомалар жұптығына сәйкес, екі доминантты аллель АА болуы керек. Осыған сәйкес ақ гүлі бар басқа ата-аналык өсімдіктің клеткаларында гомозиготалы қалпында ақ түстің рецессивті аллельдері, яғни аа болады.
Мейоз нәтижесінде әр гаметада хромосома жұбының біреуі ғана қалады, сөйтіп бір аллель А (өсімдігінде АА) немесе а (өсімдігінде аа) болады. Ұрықтану нәтижесінде будан зкго-тасында хромосома жұбы қалпына келеді және будан формула-сы дәл Мендель сипаттап жазғандай — Аа болады. Будан ор-ганизмінде жыныс клеткалары түзілуіндегі мейозда жүп хро-мосомалар ажырап әр түрлі гаметаларға кетеді, бүл кезде ген-нің бір-біреуден А немесе а аллелі бар аталық және аналық гаметалар тең мөлшерде түзіледі. Ұрықтану кезінде екі типті аталық және аналық гаметаларының қосылу мүмкіндігі бірдей, осының нәтижесінде белгілердің ажырауы. ІАА : 2Аа : Іаа тү-рінде болады.
Реципрокты шағылыстыру.Қарастырған мысалда аналық өсімдікте

| 23-сурет. Ғі буданын рецессивті ата-ана формасымен қайыра шағы-лыстыру — анализдеуші шағылысты-ру. Факторлардық белгіленуі 22-су-реттегідей. |
доминантты, аталық өсімдікте рецессивті белгі болды. Ал егер керісінше аналық өсімдікте рецессивті белгі, аталық. өсімдікте доминантты белгі болса, түқым қуалаудың сипаты өз-* гере ме? Бұл сүраққа жауап беру үшін реципрокты шағылысты-ру дегенді жүргізу қажет. Феципрокты шағылыстыру кезчіде доминантты және рецессивті белгісі бар организмдер аналық ретінде де, аталық ретінде де пайдаланылады. Кейде реципрокты шағылыстыруда тура шағылыстыру және кері шағылыстыру деген болады. Мысалы, тура шағылыстыру, ал $ аа\ &АА — кері шағылыстыру.
Көпшілік жағдайда реципрокты шағылыстырудың нәтижесі бірдей, яғни Ғх кезінде будан организмдер біркелкі, белгілі бір белгінің аталық немесе аналық организмнен келгеніне қарамай, Ғ2 кезінде белгілердің ажырауы бірдей болады. Белгілердің тұқым қуалауының хромосомалық механизміне сүйеніп, мүны оңай түсіндіруге болады.
2. ҚАЙЫРА ШАҒЫЛЫСТЫРУ
Қайыра шағылыстыру.Гибридологиялық анализ үшін буданды ата-ананың біреуімен шағылыстыруды пайдалануға болады. Гибридтің бірінші үрпағын гомозиготалы жағдайдағы белгілі бір аллельдер жүбы (доминантты немесе рецессивті) бар формамен осылайша шағылыстыруды қайыра шағылыстыру немесе беккросс деп атайды, ал ондай ұрпақты Ғв деп белгілейді. Генетикалық анализ үшін осы екі шағылыстырудың нәтижесі әр түрлі болады.
Буданның (Аа) бірінші ұрпағын Ғь доминантты аллель бо йынша гомозиготалы (АА) бастапқы формамен қайыра шағылыстырған кезде, ата-аналық өсімдіктің барлық гаметасында доминантты аллель А болады, ал буданда гаметаның екі сорты А және а болады. Сондықтан осындай гаметалардың үрықтану кезшдеп кездеисоқ үилесшдері ұрпақтағы генотип бойынша2Аа:2АА немесе 1:1 қатынасында болады, ал фенотип бойынша белгілердің ажырауы гбайқалмаиды; Ғ,, кезшде барлық есімдіктердің гүлі қара қошқыл келеді.
Анализдеуші шағылыстыру. Генетикалық анализ жасау үшін Ғ\ буданын (Да) рецессивті аллельдері бойынша гомозиготалы формамен (аа) шағылыстырудық маңызы үлкен болады. бсындай шағылыстыруды анализдеуші шағылыст-ру деп атайды (23-сурет). Бұл жағдайда рецессивті форма а аллелі бар гаметаның бір сортын ғана түзеді. Осыған бай-ланысты Ғ в кезінде буданның бірінші ұрпағында екі аллель-дің кез келген біреуінің көрінуіне мүмкіндік туады. Қарақошқыл гүлі бар есімдікті Ғ в кезінде анализдеу арқылы біз оның генотипінде ақ бояулы бір аллелі бар екенін білеміз. Сөйтіп, Ғ\ буданнан қара қошқыл бояудың аллелі, А ғана келуі мүмкін. Ғ-.9 кезіндегі екіңші фено-тип — ақ гүлді өсімдіктерде — рецессквті ата-анадан келген ақ бояудың аллелі а болады. Демек, буданнан дәл осындай рецес-сивті аллель а келуі мүмкін. Сонымен, біз мынадай қорытындыға келеміз: буданның бірінші ұрпағында бір ғана генотип — Аа бо-луы мүмкін. Оның үстіне, егер Ғ 3 кезінде доминантты және рецессивті формалардың ажырауы 1 : 1 қатынасындай жүретіні байқалса, онда мынадай қорытынды шығаруға болады: гибридте А және а аллельдері бар гаметалар тең қатынаста түзіледі.
Сонымен, Ғ в кезінде белгілердің ажырау ерекшелігіне қарап, буданның генотипін, будандар түзетін гаметалар типін және олардың арақатынасын талдауға болады. Сондықтан да будан организмді рецессивті гомозиготалы бастапқы формамен шағылыстыру анализдеуші деп аталды.
Анализдеуші шағылыстыру арқылы шыққан тегі белгісіз организмнің генотипін тексеріп анықтауға болады. Мысалы, гүлі қара қошқыл өсімдіктерді гүлі ақ әсімдіктермен шағылыстырған кезде алынған ұрпақтың бәрінің гүлі қара қошқыл болса, онда онын, генотипі АА болғаны, яғни ол гомозиготалы доминантты форма болғаны. Міне, сондықтан да анализдеуші шағылыстыру дегеніміз генетикалық анализдің аса маңызды тәсілі болып табылады.
Мендель заңының қайта ашылуы.Мендель ашқан жаңалық 1866 жылы жарияланды, бірақ оны Мендельдің замандастары түсінбеді. Отыз жыл өткеннен кейін цитологтар митоз, мейоз және ұрықтану құбылыстарын зерттеп білгеннен кейін ғана ол заңды мойындауға тура келді. Сөйтіп, Мендельдің тұқым қуа-лаушы факторларын хромосомалармен және олардың митоз бен мейоз кезіндегі ерекшеліктерімен байланыстыруға мүмкіндік туды.
1900 жылы үш ғалым —- Г. де Фриз, К- Корренс және Э. Чер-мак — әрқайсысы өз бетінше өздерінің зерттеулерінің нәтижесін баспада жариялады. Олар өздерінің зерттеулерін әр түрлі объектілерде жүргізіп ертеректе Мендель ашқан негізгі заңдылықтарды растады. Мендель әдісінің және ол анықтаған түқым куалау заңдылығының қайта ашылуы тқым қуалаушылық пен өзгергіштікті зерттеуге аса зор ұйтқы және ғылымның жаңа саласы — генетиканың жарыққа шығуына себеп болды.
Әр түрлі объектілерде алуан түрлі белгілердің тұқым қуалауын зерттеу нәтижесі тұқым қуалаудың негізгі заңдылықтары барлық өсімдіктер мен жануарлар — омыртқалы, омыртқсыз жануарлар үшін де, сол сияқты адам, микроорганизмдер үшін де ортақ екенін көрсетті. Сонымен қатар тұқым қуалауы Мендель заңына симайтын көптеген фактілер жиналды. Осыған қарамай, мендельдік зерттеу әдісі ғана осындай ауытқулардың табиғатын анықтауға және тұқым қуалаудың негізгі заңдылықтарының барлық организм үшін жалпылама ортақ екенін дәлелдеуге мүмкіндік берді.
3. ДОМИНАНТТЫЛЫҚТЫҢ ТОЛЫҚ БОЛМАУЫНДАҒЫ ТҰҚЫМ ҚУАЛАУ. ДОМИНАНТТЫЛЫҚ СИПАТЫНЫҢ ӨЗГЕРУІ
Доминанттылықтың толық болмауы.Өсімдіктер мен жануарлардың көптеген белгілеріне аралық немесе доминанттылық-тың толық болмау құбылысы тән, мүны Мендельдің өзі бұршақтьқ кейбір белгілері бойынша байқаған болатын. Доминанттылықтың толық болмауы кезінде Ғ\ бұдан да (Аа) ата-ана белгілерінің әрқайсысы толық күйінде көрінбейді, белгілер аралық түрде, ата-ананың біреуінің белгілері азды-көпті басымдау түрде керінеді, ал бұл ұрпақтың барлық особьтары өздерінің фенотипі бойынша бірдей болады.
Ақ және қызыл жемісі бар қойбүлдірген формаларын (Ғгагіа ұезса) шағылыстыру нәтижесінде алынған будан жемістерінің қызыл түс пен ақ түс аралығында қызғылт түсті болуы доминанттылықтың толық болмағанына мысал бола алады (24-сурет).
Доминанттылық толық болмаған кезде екінші ұрпақта белгілердің фенотип және генотип бойынша ажырауларының сәйкестігі байқалады, өйткені доминант гомозиготаның АА гетеро-зиготадан Аа айырмасы бар. Мысалы, біз қарастырып отырған мысалдағы Ғ2 кезінде қой бүлдірген жемістері бояуының ажырауы мынадай болады: 1 қызыл (АА) : 2 қызғылт (Аа) : 1 ақ (аа). Бүл жағдайда жемістің қызыл бояуын шартты түрде ғана
бояулы,доминантты деп, ал ақ бояуын рецессивті деп атауға болады. Сондықтан доминанттылық толық болмаған кезде кейде аллельдерді А не бояулы,доминантты деп, ал ақ бояуын рецессивті деп атауға бол анықталды. Мысалы, осындай құбылыс тауық қанаты бояуының, адам шашының бүйралығының, сол сияқты өсімдіктердің, жануарлардың және ады. Сондықтан доминанттылық толық болмаған кезде кейде аллельдерді А не а а адамның баска да көптеген морфологиялық, физиологиялық қасиеттерінің тұқым куалауында байқалады. Адамның және бірқатар үй жануарларының қан группаларын зерттеу нәтижесінде альтернативті белгілер бар екендігі жөнінде мәліметтер

|
Адамның және бірқатар үй жануарларының қан группаларын зерттеу нәтижесінде альтернативті белгілер бар екендігі жөнінде мәліметтер алынды. Олар Ғі буданында бірдей дәреже-де көрінеді, осы құбылыс кодоминанттыльщ деп аталады. Мы салы, егер аналық организмнің қан группасы А, аталық орга-низмдікі В болса, онда балалардың қан группасы АВ болады, Осы арада доминант құбылысының жоқтығына қарамай, Ғг бу-дандары бірдей болады, демек Мендельдің бірінші заңы бұл құбылыс үшін де дүрыс, яки универсал болғаны.
Доминанттылықтың мәні.Доминанттылық дегеніміз гетерозигота күйінде белгінің дамуын геннің басқару қасиеті. Бүл жағдай рецессивті аллель толық түншықтырылды және енді ол мүлде қызмет жасамайды дегенді білдіре ме? Кейбір фактілер бүл сүраққа кері жауап береді. Мысалы,қаралық доминанттылық кезінде геннің екі аллелі де қызмет жасайды. Тіпті толық доминанттылық кездің өзінде рецессивті аллельдің белгілі бір

|
өнім беретінін кездестіруге болады. Мысалы, есінек (Апііггігшт таі]из) өсімдігі гүлінің қызыл бояуы оның ашық қызғылт бояуын түншықтырады. Биохимиялық анализ мынаны көрсетті: қызыл пигмент цианидиннің ашық қызғылт пигмент пеларго-нидиннен айырмасы, қызыл пигмент-тің үшінші күйінде артық гидроксилі ОН болады (III таблица). Ол гидроксил А аллелі жасап шығаратын фермент әсерімен оның молекуласына қосылады. Мүнда рецессивті аллелі а да әрекет жасайды, бірақ оның жасап шығаратыны активті фермент емес. Осынын, нәтижесінде гидроксилдену реакциясы жүрмейді, циаыидин түзілмейді, гүлі ашық қызғылт болады. Гетерозиготада активті (А), активсіз фермент (а) түзіледі, бүнда активті фермент есе-бінен гидроксилдену реакциясы жү-реді. Сондықтан гетерозиготалы өсімдікте ақырында цианидин синтезделеді де қызыл бояу аиіық қызғылт бояуды түншықтырады.
Сонымен, доминанттылық гендерәсерінен түзілген өнімдердін. өзара әрекеттесуінің нәтижесі — доминант. ты не рецессивті аллельдер анық-тайтын белгілер түрінде көрінеді.
Доминанттылықтың өзгеруі.Доминанттылық механизмі оғіың езге-ру мүмкіндігін де қамтамасыз етеді. Шоминанттылық сыртқы орта әсер-"лерінен езгереді, орта жағдайы белгілер жұбының біреуінің дамуына себеп болады .
(Тгііі-сшп) кәдімгі жағдайда өсірген кезде қалыпты масақ доминанттылық көрсетеді, ал қысқа күн жарығында есірген кезде бүтақтанған масақ доминанттылық көрсетеді. Бірақ гибридтің жеке дамуы кезіндегі доминаттылықты өзгеруі оның генотипін өзгертпейді, сейтіп, оның ұрпағындағы белгілердің ажырауын езгертпейді. Сондықтан Ғ\ буданында қандай белгінің басым, доминант болғанына қарамай Ғ2 кезіндегі белгілердің ажырауы бірдей болады (25-сурет).
4. АЖЫРАУ ЗАҢЫН ҚАМТАМАСЫЗ ЕТЕТІН ЖАҒДАЙЛАР
Ажырау заңының көріну жағдайлары.Ажырау заңы тірі та-биғаттың басқа да заңдары сияқты тек белгілі бір жағдайда ғана көрінеді. Егер ондай жағдайлар сақталмаса, онда белгі-лердің ажырау сипаты да өзгеруі мүмкін.
Ажырау заңдылығын қамтамасыз ететін бірінші шарт — бундандардың гаметалардың барлық сорттарын түзу мүмкіндігінің бірдейлігі.
Екінші шарт — үрықтану кезінде бүкіл гаметалардың барлық үйлесу мүмкіндігінің бірдейлігі.
Үшінші шарт -— барлық генотиптер зиготаларының тіршілік қабілетінің бірдейлігі.
Төртінші шарт — организмнің даму жағдайына қарамай белгілердің толық керінуі: осындай жағдайларда ғана белгілер меидельдік ереже бойынша 3 : 1 түрінде ажырайды.
Жоғарыда аталған шарттардың біреуі сақталмаған кездегі жағдайларды нақты мысалдардан қарастырайық.
Будандар гаметалардың әр түрлі сорттарын тең түзбеген кез-дегі»ажыраудың өзгеруі.Жүгерінің бір формасы тұқымында антоциан пигментінің бар немесе жоқ болуы аллельдер жұбына тәуелді болады. Доминантты аллель А боялған, ал рецессивті аллель а боялмаған тұқым түзеді. Осы гендер бар гомологиялық хромосомалар микроспорогенездегі мейозда қалыпты ажырайды, Аа гетерозиготада микроспоралар А және а теңдей қа-тынаста түзіледі. Бірақ мегаспорогенезде хромосомалардың бі-реуі (ол 10 а деп аталады) 70% жағдайда мегаспораға енеді, ал екіншісі (10 п) дегенерацияға үшырайтын клеткаға түсе-ді. Осының нәтижесінде жұмыртқа клетканың 70%-де хромосома 10 а, тек 30%-де ғана хромосома Юя болады. • Егер Аа буданда А аллелі Юа хромосомада, а аллелі 10« хромосомада болса, өздігінен тозаңдануда гаметалардың жиілігін бірліктің бөлігі ретінде қарастыра отырып ажырау қалай болатынын есептеп шығаруға болады. Біз ұрықтаиу кезінде өзара қосылысатын аналық және аталық гаметалардың түз луін, бір-біріне тәуелсіз екі құбылысты қарастырамыз. Тәуелсіз екі құбылыстың үйлесуінің математикалық заңы былай айты-лады: бір-біріне тәуелсіз екі қүбылыстың немесе оқиғаның бір мезгілде болу ықтималдығы — олардың әрқайсысының болу ықтималдығынын, көбейтіндісіне тең. Демек, әр түрлі зигота сорттарының пайда болу ықтималдығын анықтау үшін, тиісті гаметалар жиіліктерін өзара кәбейту қажет.
Нәтижесінде генотип бойынша ажырау мынадай болады: 0,35 АА : 0,50 Аа : 0,15 аа.
Боялған дән мен боялмаған дәннің арақатынасы 3 : 1 емес, 0,85:0,15 немесе 6: 1 қатынасындай болады. Сонымен, егер бірінші шарт — будандарда гаметалардың барлық сорттарының тең ықтималды түзілуі сақталмаса, онда белгілердің ажырау сипаты езгереді.
Ұрықтанудағы гаметалардың үйлесу ықтималдығы тең бол-

маған жағдайда ажыраудың өзгеруі.Тышқандардың (Миз тизсиіиз) қысқа құйрықты болуының тұқым қуалауын мысалға альш қарастырамыз. Тышқандардың қыска құйрықты болуы доминант аллельмен А, құйрығының қалыпты болуы рецессивті аллельмен а анықталады. Егер гетерозиготалы қысқа құйрықты ұрғашы тышқанды (Аа) қалыпты құйрықты еркек тышқанмен (аа) шағылыстырса, алынған ұрпақтың жартысы қысқа қуй~ рықты (Аа), жартысы қалыпты құйрықты (аа), сөйтіп белгілердің ажырауы 1 : 1 қатынасындай болады. $ ааУ^ & Аа түрінде алып, керісінше шағылыстырған кезде, алынған ұрпақтың басым көпшілігі фенотипі бойынша қалыпты құйрықты (аа),^&е£ шағын бөлігі ғана қысқа құйрықты (Аа) болады. Мысалы, тәжірибелердің біреуінде белгілердің ажырауы 1 : 1 қатынасындай емес, 20 қалыпты құйрықты, 1 қысқа құйрықты алынды. Жүр-гізілген цитологиялық зерттеу нәтижесі еркек организмдерде (Аа) сперматогенез кезінде қалыптан ауытқу байқалмайды,. сондықтан А және а гаметалары теңдей мөлшерде түзілуі тиіс,, ал белгілердің ажырауындағы ауытқу мынаған байланысты бо луы мүмкін: ұрықтану процесінде көбінесе а жүмыртқа клетка-лары а сперматозоидпен жиі қосылады, ал А сперматозоидпен сирек қосылады. Сәйтіп, ұрықтанудағы талғаушылық деп ата-латын қүбылыс байқалады.
Зиготалардың тіршілікқабілеті бірдей болмаған кезде ажырауының өзгеруі. Англияда ірі қараның (Воз іаигиз) декстер деп аталатын тұқымы бар. Олардың дене қүрылысы өте іркілдек келеді. Осындай жануарларды өзара шағылыстырған кезде ұрпағы 2 : 1 қатынасында декстерге және қалыпты қүрылыо ты керри деп аталатын жануарға ажырайды. Белгілердің осы-лайша ажырауы декстер деп аталатын ірі қараның гетерозиготалығынан деп болжауға болады. Мұндай болжаудың дұрыс екендігін декстер мен керриді шағылыстырудың нәтижесі кәрсетті. Мұндай шағылыстыру нәтижесінде декстер мен керриге ажырау нәтижесі 1 : 1 қатынасындай болады. Демек, декстер генотипі — Аа, керри генотипі — аа. Мүндай Аа гетерозиготалы жануарларды өзара шағылыстырғанда, оның үрпағында бел-гілердің ажырауы мендельдік — 1 АА : 2 Аа : 1 аа болуы тиіс еді, оның орнына 2 Аа : 1 аа болады, ал гомозиготалы декстер АА ешқашанда байқалмайды. Ал бүзаулардың шамамен 1/4 бөлігі не эмбриональдық кезінде немесе туғаннан кейін көп ұзамай өліп қалады. Олардың өлуінің негізгі себебі сүйегінің, әсіресе бас сүйегінің кемістігінен болады. Мұндай бұзаулар — «бульдогтар» деп аталады.
Сонымен декстер типінің тұқым қуалауын мынадай түрде көрсетуге болады:
Демек, ірі қараның декстер деп аталатын типІнің дене құрылысын анықтайтын доминантты аллель, организм дамуында тұқым қуалайтын рецессивті кемістікпен байланысты және го-мозиготалы жағдайда өлімге шыратады, яғни екі белгіні анықтайды.
Сол сияқты екі әсер үшін де рецессивті болатын гендер де белгілі. Қөптеген өсімдіктер арасында хлорофилл түзе алмайтын және өздері тұқым қуалайтын ақ түсті альбинос формалар белгілі. Мүндай ақ түсті өскіндер тіршілік ете алмайды. Мысалы, бидайда көк жасыл бояу доминантты да, бояусыз болу ре-цессивті. Егер гетерозиготалы жасыл түсті Аа өсімдіктерді ша-ғылыстырса, олардың үрпағы 1 ЛА : 2 Аа : 1 аа қатынасындаажырауы тиіс немесе 3 көк жасыл: 1 түссіз өсімдік пайда бола-ды. Ал хлорофилі жоқ аа өскіндер еледі, егер ересек есімдіктер бояуын ескеретін болсақ, онда ажырау тіпті табылмайды.
Белгілердің көріну сипатына қарай ажыраудың өзгеруі.Жүгері өскінінің бояуын бір жұп аллель анықтайды: доминантты аллель А — жасыл, рецессивті аллель а — сары бояу береді. Сары бояу біршама төменгі суық температурада пайда болады. Сондықтан да егер ескіндер көктемдегі суық кезде өсіп шықса, онда будан өсімдіктің Аа ұрпақтары ажырап 3 жасыл: 1 сары өсімдік береді. Ғ2 будандар жаздағы жоғары температура жағдайында есіп шықса, онда барлық өскіндер, олардың ішінде рецессивті гомозиготалардың өзі аа жасыл болып өседі және мұндай жағдайда ажырау болмайды (26-сурет).
Ажыраудың классикалық типі 3 : 1 қатынасының бұзылуы Мендель анықтаған заңдылықтарға нүқсан келтіре алмайды. Ондай жағдайлар әр түрлі заңды немесе кездейсоқ факторлар-дың әсер етуінен болатынын көрсетеді және белгілердің тұқым қуалауын зерттеп білу үшін.бұл әдістің маңызын айқындай түседі.
5. АЖЫРАУДЫҢ СТАТИСТИКАЛЫҚ СИПАТЫ
Ажыраудың ықтималдық сипаты.Будандар үрпағында ажырауын қамтамасыз ететін механизм мейоз болып табылады. Гамета түзілу кезінде гомологиялық хромосомалардың заңды ажырауын мейоз қамтамасыз етеді. Сөйтіп, ажырау гаплоидты гаметаларда хромосомалар және гендер деңгейінде жүзеге асады, ал белгілердің ажырау деңгейінде диплоидты организмдерге

^и аа
26-сурет. Өніп шығу кезіндегі температураға байланысты жүгерінің өскіні бояуының тұқым қуалауы:
А— жасыл түсті; а— сары тусті өскіндер, ондай бояу төменгі температура кезінде байқалады.
анализ жасалады. Бүл екі процесс аралығында кеп уақыт өтеді, бұл аралықта ұрықтану болады, организм дамуының күрделі процестері жүреді. Осы кезде гаметаларға және зиготаларға және дамып келе жатқан организмге біріне-бірі тәуелсіз кәптеген жағдайлар әсер етеді. Дәл осы кезде белгілердің ажырап көрінуіне жоғарыда атап өткен факторлардың төрт тобы әсер ете алады.
Егер ажырау процесінің негізіне биологиялық механизмдер жатса сол механизмдердің көрінісі, яғни бізге байқалатын белгілердің ажырауы кездейсоқ немесе статистикалық сипатты болады.
Жоғарыда аталған төрт шарт белгілердің ажырауына кездейсоқтық элементін туғызады, сондықтан оны талдау кезінде арнаулы математикалық әдістер қолдану қажет болады. Бұл әдістер ажыраудың теория жүзінде болуға тиісті ажыраулардан (3 : 1, 1 : 1 т. с. с.) ауытқуы кездейсоқ емес пе, ажырауды бұзу-шы факторлардың заңды әсерінен емес пе (мысалы, белгілі бір генотипі бар зиготалардың өліп қалуы) немесе ол анализ жасалатын материалдың аздығынан емес пе — осылардын, бәрін шешуге математикалық әдіс мүмкіндік береді. Ықтималдық теориясы бойынша нақты алынған мәліметтің теория жүзінде күтілген мәліметтен ауыткуы көбінесе зерттелген материалдьщ аз алынғандығынан, кездейсоқ себептер әсерінен болады.
| 12 буданды өсімдіктер ұрпағындағы ажырау |

|
1-таблица Мендельдің жүмысынан келтірілген 1-таблицадан көрінгендей, 12 өсімдік ұрпағы дәндерінің бояулары бойынша ажырап айрылуы аз мөлшерде алған кезде өте түрленіп, алшақтап кеткені байқалады.
Ажырау заңдылығына алынған өсімдік санының әсерін осы мысалдан көруге болады. Талдау үшін № 12 өсімдік ұрпағын алып қарастырайық, бұл мысалда ажырау 3:1 қатынасынан көп ауытқуы байқалады. Бүл өсімдіктің дәнін барлық есімдіктен алынған дәнмен салыстырып көреміз.
Ауытқудың кездейсоқты-ғына статистикалық баға беру үшін %2 (хи-квадрат) әдісін қол-данады1.
33 түқымнан тұратын шағын іріктеуде ажыраудың көрінісі күтілген 3 : 1 қатынасымен салыстырғанда кездейсоқ және өте шамалы ғана (Р<С0,01), яғни бүл қарастырылған іріктеуде 3 : 1 жоқ. Бұл мысалдағы жасыл түсті дәннің аздығы ескерілмеген бір себепке байланысты болуы мүмкін. Айталық: негізінен жа-сыл дәндер бар бүршақ қындары түсіп қалады немесе ондай дәндері зиянкестердің әсерінен қүртылған болуы керек. Тұқым1 Ватти Қ. В., Т и х о м и р о в а М. М. Руководство к практическим занятиям по генетике. М., Просвещение, 1979.саны аз (33) кезде әр кластьщ 3 : 1 (7,2) қатынасынан ауытқуы кездейсоқ болмайды. 550 тұқымнан тұратын іріктеуде де дәл сондай ауытқу (5,5) кездейсоқ (Р^>0,05) болады, яғни мұнда белгілердің ажырауы 3 : 1 болады.
6. ГАМЕТАЛЫҚ АЖЫРАУ ЖӘНЕ ТЕТРАДАЛЫҚ АНАЛИЗ
Ажырау заңдылығын тұтас организм белгілері бойынша ғана емес, сол сияқты мейоз өнімі болып табылатын гаметалар белгілері бойынша да анықтауға болады.
Төменгі сатыдағы өсімдіктердегІ гаметалық ажырау.Кейбір төменгі сатыдағы организмдерде ажырауға жекеленген тет-рада деңгейінде анализ жасауға болады.
Саңырауқұлақтар, мүктер және балдырлардың көпшілігінде ұрықтанудан кейін бірден мейоз басталады. Сондықтан ондай организмдерде диплоидты фазаның (зиготаның) ұзақтығы өте

аз, ал организмдердің тіршілік цик-лінде гаплоидты фаза ұзаққа созылады, осы фаза кезінде әр аллельдің доминантты және рецессивті әсері көрінеді. Төменгі сатыдағы организмдердің мұндай еоекшелігі гаме-талардың және олардан дамып жетілген гаплоидты особьтарының ж-ке белгілерінің ажырауына анализ жасауға мүмкіндік береді.
| А й Ашытқы Зассһа-колония:ларыньщ |
| 27-сурет. готусез бояуыньщ тұқым қуалауына жасалған тетрадты ана-лиз: А— ак,; а— ңызыл. |
Тетрадалық анализ.Тетрадалық анализ деп аталған осы әдіс мыңаны дәлелдеуге мүмкіндік берді. Мен-дельдік ажырау мейоз механизмімен негізделеді жәие ол статистикалық емес, биологиялық заңдылыққа ба-ғынады. Тетрадалық анализ әсіресе төменгі сатыдағы саңырауқүлақтарда: зең саңырауқүлағында (Меигозрога сгазза) және ашытқы саңырауқұлақта (8ассһаготусе5 сегеуізіае) табысты пайдаланылды. Осы саңырауқүлақтарда мейоз нәтижесінде төрт аскоспора, яғни бір қап-шықтағы — аскадағы споралар түзіледі. Споралардың асқадағы орналасуы әр түрлі болуы мүмкін: не линия бойына, немесе бөліну (нейроспораларда), не секториальды (ашытқыларда) осіне орналасады. Микроманипулятор кемегімен спо-ралардын, әрқайсысын бөліп алуға, еөйтіп олардың көбеюіне мүмкіндік жасауға болады. Ашытқылардағ тетрадалық анализді мысалға алып көрейік. Саңырауқүлақтың бұл түрінде қызыл және ақ колониялар кездеседі (27-сурет). Мұн-дай альтернативті белгілер аллельдер жұбымен анықталады А— ақ түсті, а — қызыл түсті колониялар. Гаметалар (споралар) қо-сылуы кезінде диплоидты зигота Аа түзіледі. Көп ұзамай дипло-идты зигота мейозға кіріседі, соның нәтижесінде бір аскада гаплоидты споралардың тетрадаеы түзіледі. Содан кейін асканы жарып, әр спораны жеке алады да, қоректік ортаға орналастырады. Гаплоидты төрт клетканың әрқайсысы белініп көбейеді, сөйтіп төрт колония түзіледі. Олардың екеуі ақ, екеуі қызыл түсті, яғни 1 А: 1 а қатынасында болады. Бұдан басқа да белгі-лер жұбының әрқайсысында осындай қүбылыс байқалады.
Тетрадалық анализден мынадай жағдай айқындалады: моногибридті шағылыстыру кезінде гендердің ажырауы мейоздық бөлінудің нәтижесі болып табылады.
7. ТҰРАҚСЫЗ ТИПТІ ЖЫНЫСТЫ ЖӘНЕ ЖЫНЫССЫЗ КӨБЕЮ ДЕП ТҰҚЫМ ҚУАЛАУШЫЛЫҚ ЕРЕКШЕЛІГІ