ПОЛОВОЙ ХРОМАТИН
Половой хроматин — это структура интерфазного ядра соматических клеток млекопитающих, образованная в результате гетерохроматинизации половых хромосом. Половой хроматин является формой факультативного гетерохроматина, т.к. отличается в клетках разного пола и по происхождению:
- хроматин X является результатом гетерохроматинизации одной из двух хромосом у женщин;
- хроматин Y образуется в результате гетерохроматинизации 2/3 дистального плеча
хромосомы Y у мужчин.
Изучение полового хроматина позволяет установить генетический пол (XX или XY) и нарушения половых хромосом.
ИНАКТИВАЦИЯ ХРОМОСОМЫX
Половые хромосомы (по Susumu Ohno, 1967) произошли от аутосом, которые в ходе эволюции дифференцировались генетически и морфологически, образовав хромосомы X и Y. Хромосома Y является результатом длительной прогрессивной „специализации", в процессе которой были сохранены гены дифференцировки пола и утеряны почти все аутосомные гены, а размеры хромосомы стали намного меньше. Хромосома X сохранила не только исходную форму, но также и большинство генов, как аутосомных, так и связанных с половой дифференцировкой. Различия в половых хромосомах не случайны, а имеют важное биологическое значение, так как они:
- препятствуют обмену генами между хромосомами X и Y в мейозе и обеспечивают сохранение в чистом виде половых детерминант каждой из половых хромосом;
- обеспечивают образование при оплодотворении зигот разного пола: XX или XY.
Половые хромосомы(гоносомы, гетеросомы) отличаются по структуре (длина, положение центромеры, количество гетерохроматина) и по содержанию генов.
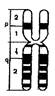
 Хромосома X- средняя метацентрическая хромосома (группа С); представлена в соматических клетках обоих полов: в двойном экземпляре в женском кариотипе - 46,ХХ и в единственном экземпляре - в кариотипе мужчин - 46,XY. В половых клетках хромосома X представлена следующим образом: в одном экземпляре во всех яйцеклетках и у 50% сперматозоидов. Хромосома X богата эухроматиновыми участками и содержит 1336 генов, среди которых:
Хромосома X- средняя метацентрическая хромосома (группа С); представлена в соматических клетках обоих полов: в двойном экземпляре в женском кариотипе - 46,ХХ и в единственном экземпляре - в кариотипе мужчин - 46,XY. В половых клетках хромосома X представлена следующим образом: в одном экземпляре во всех яйцеклетках и у 50% сперматозоидов. Хромосома X богата эухроматиновыми участками и содержит 1336 генов, среди которых:
■ структурные соматические гены (например, гены групп крови Xg, факторов свертывания крови VIII и IX, фермента 6-фосфатдегидрогеназы, цветного зрения и др.);
■ регуляторные гены феминизации,
■ структурные гены феминизации,
■ структурные гены маскулинизации.
Хромосома Y-мелкая акроцентрическая хромосома (группа G); 2/3 дистального плеча q представлены гетерохроматином в генетически неактивном состоянии. Хросома Y представлена одним экземпляром во всех соматических клетках индивидов мужского пола с кариотипом 46ХY и у 50% сперматозоидов. Она содержит около 300 генов среди которых:
■ регуляторные гены маскулинизации (SRY=Tdf)
■ гены обеспечивающие фертильность (AZF1, AZF2)
■ структурные соматические гены (фактор контроля роста зубов, рецептор интерлейкина)
■ псевдогены
Так как в женском кариотипе две хромосомы X, а у мужчин - только одна, то логично предположить, что в клетках женского организма должно быть в два раза больше конечных продуктов генов, локализованных в хромосоме X, чем в клетках мужчин. Однако, в реальности это не так, так как одна из хромосом X у женщин (в норме) или у индивидуумов с дополнительной хромосомой X (при патологии) инактивируется. В результате активной у обоих полов остается лишь одна хромосома X. Данное явление называется компенсацией сцепленных с X хромосомой генов
Гипотеза компенсации была сформулирована М. Лайонв 1961 году и включает три основных положения:
I. В соматических клетках млекопитающих активной является одна хромосома X, в то время как другая - инактивируется путем гетерохроматинизации с образованием тельца Барра, различимого в интерфазном ядре; инактивированная хромосома X реплицируется в конце фазы S.
II. Инактивация происходит на 16-й день эмбрионального развития, когда эмбрион состоит из ~3000-4000 клеток. До этого момента в каждой клетке женского эмбриона функционируют обе хромосомы X, т.е. вырабатывается вдвое больше, чем у мужских эмбрионов,- мРНК и ферментов, закодированных генами Х-хромосомы; вследствие этого, эмбрионы 46,ХХ,я 46,XY биохимически и функционально отличаются. Инактивация одной Х-хромосомы остается в дальнейшем неизменной у всех потомков данной клетки.
III. Процесс инактивации носит случайный характер, поэтому в половине клеток активной сохраняется материнская хромосома X, а в другой половине клеток активной остается отцовская хромосома X.